第二十七章 礁和礁相
生物礁无论在地史时期还是现代,都有广泛的分布,同时,它也是碳酸盐沉积中的一种重要的含油气沉积类型。在国内外都已发现了许多生物礁油气田,如加拿大泥盆系的礁油气田,美国五大湖区早古生代的礁油气田,前苏联泥盆系、石炭系、二叠系中的礁油气田,我国二叠系、三叠系和古近一新近系的礁油气田等。
由于生物礁是良好的油气圈闭,因此我国石油地质学家和沉积学家十分重视对古代和现代生物礁的研究,已先后在陕南、湘西寒武系,浙西、陕南奥陶系,川西北、陕南志留系,滇、黔、桂地区泥盆系、二叠系,川西北地区泥盆系、二叠系、三叠系和川东地区二叠系,湖南、赣西北二叠系等发现了大量的生物礁沉积体,同时在古近一新近系海、陆相地层中也分别发现了礁,为我国南方和海上油气勘探作出了巨大贡献。
第一节 礁沉积环境特点及其沉积作用
一、礁的概念、基本特征及分类
(一) 生物礁的概念
礁 ( reef),这一术语来源于挪威语 rif,其含义为脊。远在19世纪中期,达尔文就对现代礁有所认识。其后不久,一些地质学家把它应用到古代岩石中,从此开始了古代礁的研究。最初,研究古代礁的人对礁的概念局限于灰质骨骼的原地堆积。然而,古代礁能够反映现代礁的特征、生物的生态、气候、环境以及地质背景的资料都是有限的。因此,一些地质学家在研究古代礁时,只能根据地质时代中保存下来的有限的资料来认识它,并且几乎都是从生物造成的海底地形上的特点来讨论礁的存在与否。这样,除了一些真正的礁外,常常把一些因海流作用造成的一些异地介壳堆积、鲕粒丘、石灰岩的残山,甚至一些砂页岩与石灰岩的相变也看成是礁,这样,礁这一术语便产生了某些混乱。
广大研究者对礁大体有两类理解,即邓哈姆 (1970) 提出的礁的双重概念:狭义礁和广义礁。
狭义的礁即所谓生态礁,它是由造礁生物原地生长而经常迎浪营建起来的; 具凸起的地貌; 具造礁生物骨架或只见造礁生物原地生长的痕迹; 分布范围从海平面到200m以下水深,视造礁生物种类不同,有的水深可达400~500m。
广义的礁实际上是指生物礁是主要由生物和生物作用 (包括宏观生物作用以及微观生物作用) 所形成的,具有地貌特征的碳酸盐岩体。这是目前较流行的一个广义定义,包括了所有由生物和生物作用成因的碳酸盐岩体 (范嘉松,1988; 吴亚生等,1991)。
(二) 礁的基本特征
礁主要由礁核和礁翼组成。在一些群礁复合体中,礁间沉积也与礁的发展有密切关系(图27-1)。
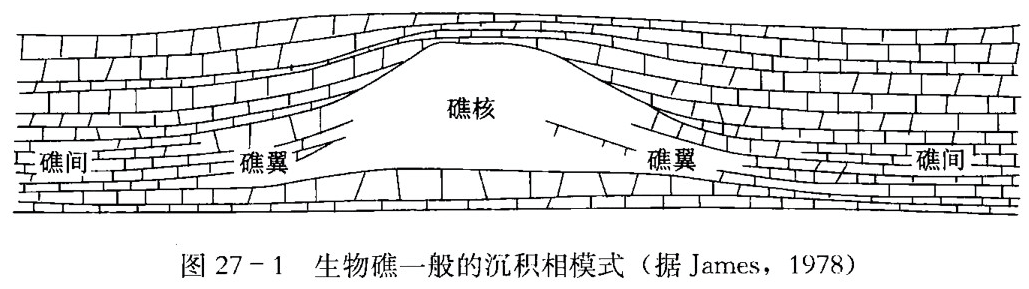
- 礁核
礁核是指礁体中能够抵抗波浪作用的部分,是礁的主体。它主要由原地堆积的生物岩或粘结岩组成。其中生物的含量很高,主要是造礁生物,还有一些附礁生物。这些造礁生物有时保存有原地的生长骨架,骨架间常有礁的破碎物充填。其原生孔洞不是被内部沉积物充填,就是被早期纤状文石和镁方解石胶结。
- 礁翼
礁翼通常是指礁相与非礁相呈指状交错过渡的那部分礁体。礁体迎风的一侧称礁前,背风一侧称为礁后。在一些盆地或潟湖的斑礁或塔礁中,由于未受到方向性风浪作用的影响,礁前与礁后极为相似,所以礁翼就分不出礁前和礁后。
礁前处于迎风一侧,在风浪冲击下,礁碎屑顺着礁前缘的陡坡堆积形成的岩石一般称礁前塌积岩或礁前礁砾岩。这些礁碎屑大都未被磨圆且分选差。通常坡前的礁屑与灰泥混积,向盆地方向砂屑增加,砾屑减少,它们与正常海的盆地相泥质沉积物呈指状接触。
礁后沉积多由分选较好的砂屑石灰岩组成,胶结物多为亮晶方解石。与礁前相比,其生物门类和种属大为减少。
- 礁间
在一些群礁复合体中,礁与礁之间的沉积物和生物组分与礁的发展有极其密切的关系。在海侵的情况下,群礁一般是发展的,在礁间可以出现正常的海相碳酸盐沉积; 当海退时,群礁的发展受到抑制,礁间可出现一些潟湖相的沉积。有时礁间也可以包括在礁组合的复合体中。
(三) 礁的分类
关于礁的类型,主要是根据其形态和地理位置来划分 (图27–2)。
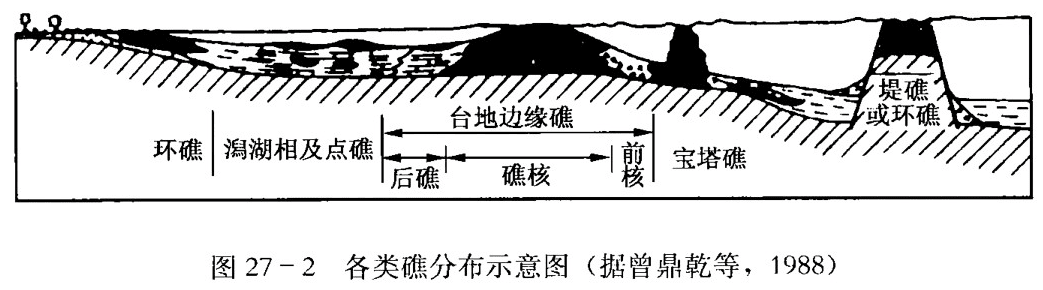
按形态可将礁分为以下类型:
(1) 点礁,也称斑点礁。礁体近似圆形,或呈不规则状,是在潟湖或外滨海底较小隆起上形成的孤立小礁体。现代海洋中,点礁主要分布在大陆架海域浪基面以上,并止于海面。实际上,常用点礁指示大陆架位置,如加拿大泥盆纪利迪尤克礁。
(2) 宝塔礁,也称尖柱礁和孤礁。形似锥状或者陡侧向上变尖的丘状,由成礁期海底持续下降而成,多出现于深水带。
(3) 马蹄形礁,也称新月形礁。向风面一侧礁体发育,背风一侧不发育,触体凸面迎风,多分布于开阔海盆中,如美国二叠纪马蹄形礁。
(4) 环礁,礁体围绕海底较大隆起的边缘生长,连接成环状,中央带凹下成潟湖,多出现于外滨广海中。现代太平洋、印度洋以及我国南海均有发育,古代礁如墨西哥白垩纪的环礁。
(5) 丘礁,孤立地分布,近似半球状,由波基面以下较深水碳酸盐堆积而成。圆丘礁或宝塔礁用来指示大陆架边缘或盆地内的单元岩隆。
(6) 层状礁,也称带状礁和滩礁。分布面积较大,礁高度不大,多分布于碳酸盐岩台地上,相当于前述生物层礁。
按礁体的地理位置又可分为:
(1) 岸礁,也称边礁、镶边礁和裙礁。紧靠海岸生长,顶平,由于向海岸一侧斜坡常很陡,故发育陡峭的海岸峭壁或陡坡,呈曲线状。有时在边缘礁与岸之间有一小的平底水道相隔,水道逐渐加宽便可发育成堡礁。现代最长的边缘礁在红海沿岸,长约2700km以上,向海一侧伸入海下36m深。
(2) 堡礁,也称堤礁、堤岛礁或障壁礁。堡礁多在平缓的海岸生长,离海岸有一定距离。平面上礁多呈曲线状,平行海岸分布,形成的堤礁与陆地之间有潟湖相隔。有时堤礁不止一排,按生物不同生态 (或生长深度不同) 或其他原因可有多排堡礁出现。现在世界上最大的堡礁是澳大利亚东北岸的大堡礁,长达2000km,向岸外延伸达50~145km。古代最大的堡礁是美国新墨西哥州东南部和得克萨斯州西部二叠纪盆地的船长礁,厚达360m以上,长达644km,现已在埋藏的地下部分找到了油气藏。
(3) 边缘礁,也和堤礁一样与盆地深度的剧烈变化有关,但远离海岸分布。尽管礁后广阔的水域较浅并比较隔绝,但礁后沉积为正常海水的碳酸盐岩层。这种类型的现代礁见于巴哈马群岛,印度洋有恰果斯群岛礁。古代有前苏联乌拉尔——伏尔加区的上泥盆统多内昔礁,阿尔兰—迪尤尔提尤利礁等。
二、礁的形成及生物造礁作用
(一) 礁的形成条件
一切礁和有机建造的形成,都与发育繁盛的、能分泌石灰质的动植物有关。因此,产生礁和有机建造的最重要的条件之一,就是要有能使礁生物群落中的生物蓬勃发展的、合适的生态条件。
在现代热带地区的碳酸盐台地上,常有珊瑚礁的分布。造礁生物主要是珊瑚和珊瑚藻(红藻),影响珊瑚生长和珊瑚礁发育的因素是水温、盐度、水深、浊度(或透明度)、溶解的气体、底质以及波浪和水流等。此外,一些海洋生物对生物礁的发育也有很大影响。
大多数的珊瑚礁一般局限在热带和亚热带,大致在南、北纬30°的范围内分布。珊瑚在现代海洋中有造礁型和非造礁型两种类型。造礁型珊瑚,在水温23~27℃的环境里生长最佳。一般说来,在冬季温度下降到18℃以下,或夏季温度上升到30℃以上的地区,造礁珊瑚就不能顺利地生长。通常,造礁型珊瑚比非造礁珊瑚的钙化速度要快。
一般认为适合珊瑚生长的海水含盐度为30‰~40‰。强烈的蒸发作用 (如波斯湾) 以及大陆径流的注入 (如海南岛的一些海湾),均会影响珊瑚种属的分布和礁的发育。
阳光是珊瑚生长的必要条件之一。珊瑚只能生长在深度不大、透明度良好的海域里。大多数的珊瑚在特别清澈的海水中可以生活在70~80m的深度内。但在一般条件下,50~60m的深度已是珊瑚生长的极限,以小于50m为宜。海水的透明度主要受陆源物质供给的影响。
波浪、海流以及风对珊瑚礁的影响也较大。它们既可以给珊瑚输送生长所需要的氧和食料,也会破坏珊瑚礁。
(二) 生物造礁作用
从生态的观点看,一般把礁的生物划分为4种类型,即礁骨架建设者、礁骨架粘结者、礁骨架居住者以及礁骨架保护者。这些具有不同功能的各种生物形成了一个对立和统一的总体。
目前,已知有许多生物 (层孔虫、苔藓虫等) 可作为积极的造礁生物,具有双重作用:在礁核,这些生物形成树枝状群体 (即普通的建架生物);在礁缘,这些生物群体获得板状蔓延形态,并可以划入胶结生物中。
现在生长着的礁一般都有丰富的原生骨架和次生骨架的建造生物。它们是珊瑚、钙质红藻、苔藓虫、牡蛎、结壳的有孔虫、蛇螺类、腹足类、龙介以及海绵,还有大量的各种各样的生物骨骼和软体动物堆积或固着在骨架上。固着在骨架上的生物有钙质绿藻 (仙掌藻)、柳珊瑚以及某些双壳类生物。自由生活在礁中的生物有腹足类、棘皮类、蛇尾类以及有孔虫。这些生物虽然不能构成坚固的骨架,但它们可以提供礁中的沉积物来源。此外,礁中还有一些钻孔生物、某些双壳类、海绵、蠕虫以及藻类。
礁群落中的生物按深度、光线、波能、淤积的条件作带状分布,同一类生物的不同种也有不同的分布范围。这种变化在巴巴多斯的更新世和现代礁中是很清楚的 (图27–3)。在礁前上部中等水深处 (5~15m),生物分异度高。
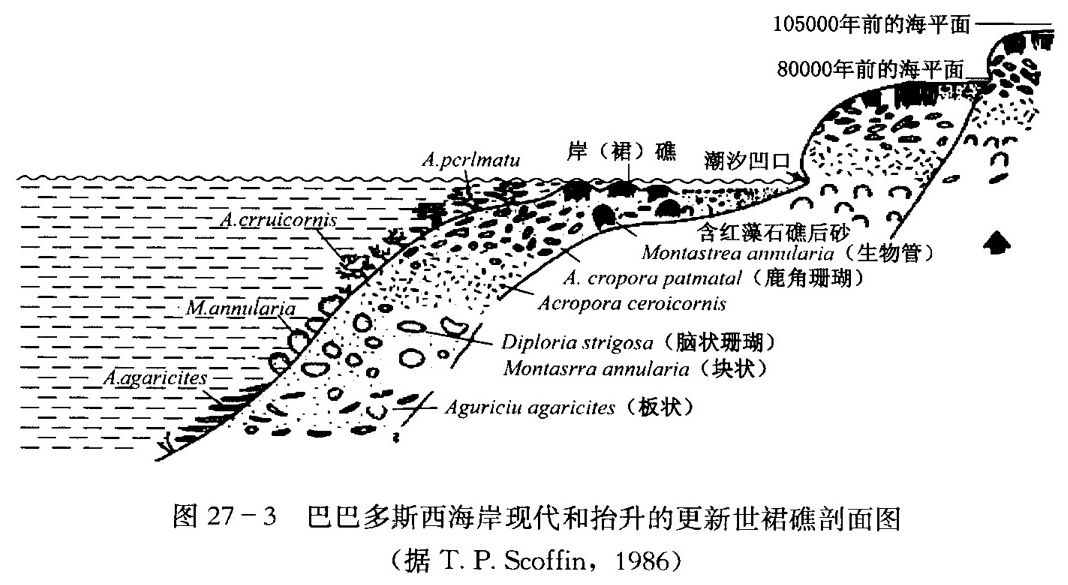
造礁生物的形态与其生长环境有密切关系,尤其是其外形和礁生长处的波浪、水流作用能量之间的消长关系,可以作为我们现今判断古代礁沉积相的重要依据。根据对岩石记录中的生物与周围沉积物之间相互关系的许多观察,再结合对现代珊瑚和热带礁分布所作的研究,可以作出关于生物外形和环境的某些概括,这对礁的相分析是很有用的(表27-1)。
表27-1 造礁生物的生长形状及其最常出现的环境类型 (据 James,1978)
| 生 长 形 状 | 环 境 | |
|---|---|---|
| 波浪能量 | 沉积作用 | |
| 纤细的、分枝的 | 低 | 高 |
| 薄的、脆弱的、平板状的 | 低 | 低 |
| 球状的、球基圆柱状的 | 中等 | 高 |
| 强壮的、树枝状的 | 中等—高 | 中等 |
| 半球状的、穹状的、不规则的、块状的 | 中等—高 | 低 |
| 结壳状的* | 强烈 | 低 |
| 薄板状的* | 中等 | 低 |
*两者在岩石记录中难于区别,但它们表示的沉积环境不同。
生物礁是生物和环境斗争的产物,伴随生物礁的生长发育,生物在其中起着决定性作用。例如,生物可以直接为礁体建造提供骨骼物质,也可以固定胶结其他生物化石; 可以改变礁体中生物生活条件,促进礁的生长,又可促使碳酸盐物质发生生物化学沉淀,对海洋礁体的生长起到积极的作用; 而同时生物又对正在建造中的礁体进行锉刮、钻孔、啃食等,破坏礁体的生长,起到消极的作用。
概括起来,生物有5种作用形式:
(1) 骨架式。造架生物 (如珊瑚等) 死亡后仍保留其生态条件,作为礁体拓展的基本格架,这种生物在礁体中起着重要作用。
(2) 障积式。如海底的海藻,当海流经过时,生物可阻碍海流中的泥晶物质而沉淀成岩。
(3) 粘结式。如层孔虫,能把海底生物碎屑覆盖起来快速粘结成岩,起到加固作用,以抵抗波浪的强烈破坏。
(4) 附着式。藻类等可以附着在骨架生物上造成结壳,起到加固作用。
(5) 胶结式。藻类生长在洞穴或孔隙内,产生胶结作用,同时起到加固作用,有许多碳酸盐颗粒都是被海藻胶结成岩的。
大部分生物礁都是这几种生物作用综合形成的,很少是某种单独的作用造岩。
总而言之,生物在礁体中可以造粒、造灰泥、造岩。
(三) 生物礁的形成
同自然界的其他事物一样,礁也有它的发生、发展和消亡的过程。在海侵过程中海平面上升的幅度太快,海水变深,或海退过程中海平面下降得太快,海水变浅,盐度增加,以及其他因素等,都会中止生物礁的发育。应当怎样来了解礁的发育过程和它们的地质背景呢? 礁的沉积物特征、礁的微相以及造礁生物群落的演替现象是揭示礁发育过程的重要途径。
生物礁是造礁生物和沉积物的镶嵌体,但是生物起着主导作用,这类生物必须具有骨架。例如珊瑚,在适宜的环境中向上生长,并会受到大群锉刮动物、钻孔动物的不断破坏,另一些生长迅速而生命较短的、附着的钙质底栖生物造成了丰富的沉积物,这是一个自然而微妙的平衡 (图27-4)。
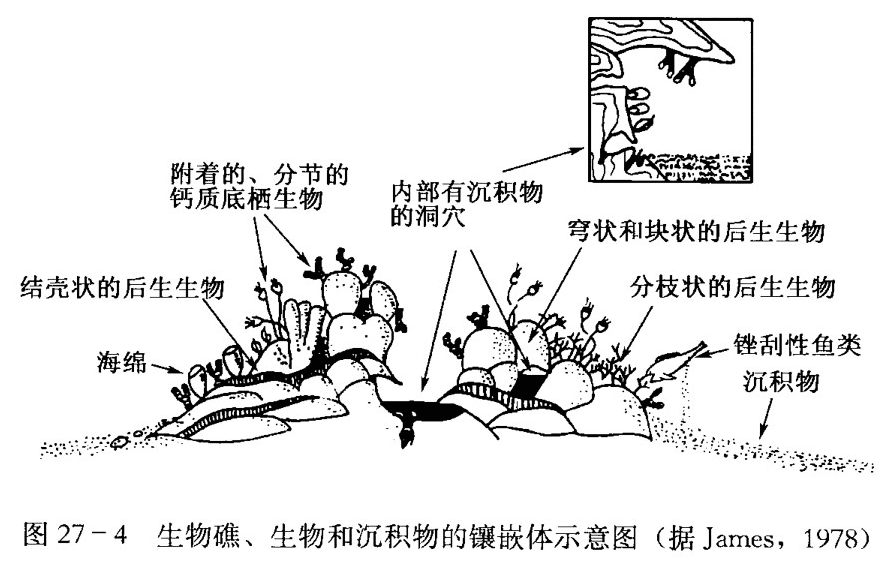
许多古代礁的研究表明, 生物存在着生态演替 ( Lonenstan, 1950; 陈源仁, 1992), 随着礁的生长,一种造礁生物群落被另一种替代了。K. L沃克和L. R艾伯斯塔特 ( Walker和 Aberstate, 1975) 关于从早奥陶世到晚白垩世礁的分布情况的综合研究表明, 在整个古生代和中生代期间的礁中,存在有以上相似的群落演替,现代礁也同样可以看到这种群落的演替。
在大多数情况下,能够辨出礁的生长有定殖阶段、拓殖阶段、泛殖阶段和统殖阶段等4个独立阶段。每个阶段都有其特点,这表现在石灰岩类型、造礁生物的相对多样性及其生长形态等方面 (图27–5)。

(1) 定殖阶段:是指生物碎屑堆积并开始被其他生物固定形成地貌高地。在古生界和中生界,最常见的是由有柄亚门或棘皮动物碎片组成的一系列浅滩或骨骼灰质砂的堆积体,新生界则由钙质绿藻的板片组成。这些沉积物的表面繁殖着藻类 (钙质绿藻)、植物(海草) 或者动物 (有柄亚门),它们围着底层,使其联结和固定下来,随后星星散散的枝状藻类、苔藓虫、珊瑚虫、软的海绵和其他后生生物就开始在定殖的生物之间生长起来。
(2) 拓殖阶段:这个单元同整个礁的构造相比,厚度比较薄,反映造礁后生物的初期繁殖。此阶段通常以生物种很少为特征,岩石形状有时是成丛的枝状。在新生代的礁中,此阶段所有珊瑚的一个共同特点是它们能够摆脱沉积物而洗净珊瑚虫,因此它们在沉积作用强烈的地区也能够生长。枝状的生长形式造成了许多较小的亚环境,形成了礁生态系统的第一阶段。这个阶段常可见层状晶洞构造。
(3) 泛殖阶段:这个阶段是礁体的主要构成时期,也是礁体向上生长最显著的时期,侧部相也发育起来,主要造礁生物的种属多,并且可以看到多种多样的生长习性。随着生物形态的增多,以及形成格架的和起粘结作用的种属数目的扩大,栖居空间 (即表面洞穴等) 也相应增多,导致产生碎屑的生物的多样化。
(4) 统殖阶段:礁体生长至这个阶段,变化常较突然。最普遍的岩性是石灰岩,并以只具有一种生长习性的 (一般是结壳的到纹层状的) 少数几个种属的生物占统治地位。在这个阶段,大多数礁受拍岸浪的影响,形成碎块灰岩层。
造成上述生态演替的原因还有争论,概括起来有3种看法:(1) 由于礁向海面和更加动荡的水域生长,生物的深水群落逐渐被浅水群落所代替。(2) 随着生物逐渐改变底层并改变水流通道而造成的一种自然演替。(3) 这种构造生长时,水的动荡性逐渐增高。由此可见,生态演替最根本的原因,还在于礁体本身的生长与生物群落两者构成了礁体不断演化的动力。
三、礁形成的控制因素
生物礁就是通过骨骼生物的生长,沉积物的充填,结壳生物的粘结等不断增生而成的。大部分生物礁内的沉积物是由分节 (海百合、钙质绿藻) 或不分节的 (瓣鳃类、腕足类、有孔虫等) 生物死亡后解体而构成的。其余的沉积物则是由侵蚀礁的各种生物产生的,如钻孔动物 (蠕虫、海绵、瓣鳃类) 产生灰质软泥; 啃食礁的表层的锉刮动物 (海胆、鱼等) 产生大量灰质砂和粉砂。
一般没有单一生物形成的生物礁。在生长条件适宜,即当食物供应充足,化学和物理条件最有利时,随着造礁群落的演替,便会出现多样的生物类群和生长形式。
影响现代造礁群落和化石造礁群落的因素有:(1) 温度和盐度; (2) 海浪和其他水动力作用; (3) 透光性; (4) 沉积作用; (5) 基底沉降与生物建礁和沉积速率的消长均衡关系。一般来说,水体温暖、水体清澈、水体动荡、水体较浅、陆源沉积较少、海平面升降或基底沉降与生物建礁速率均衡是利于生物礁生产发育的。
第二节 礁相和礁复合体沉积模式
礁建造的典型特征,就是各类岩石和化石群落在礁块中有规律的分布,从而造成生物礁体岩相上的明显分带性,为礁相带的划分和模式的建立奠定了基本轮廓。
礁复合体或礁组合是生物礁的不同相的总称,凡是与礁发育有关的相都应概括在礁复合体中。这样,除了骨架相、礁顶相等以外,礁后潟湖、礁前斜坡以及塌积岩也都被包括在礁复合体中 (图27-6,表27-2)。因为这些环境的产生都与礁的发展有联系,并且其沉积物的主要来源也是礁。
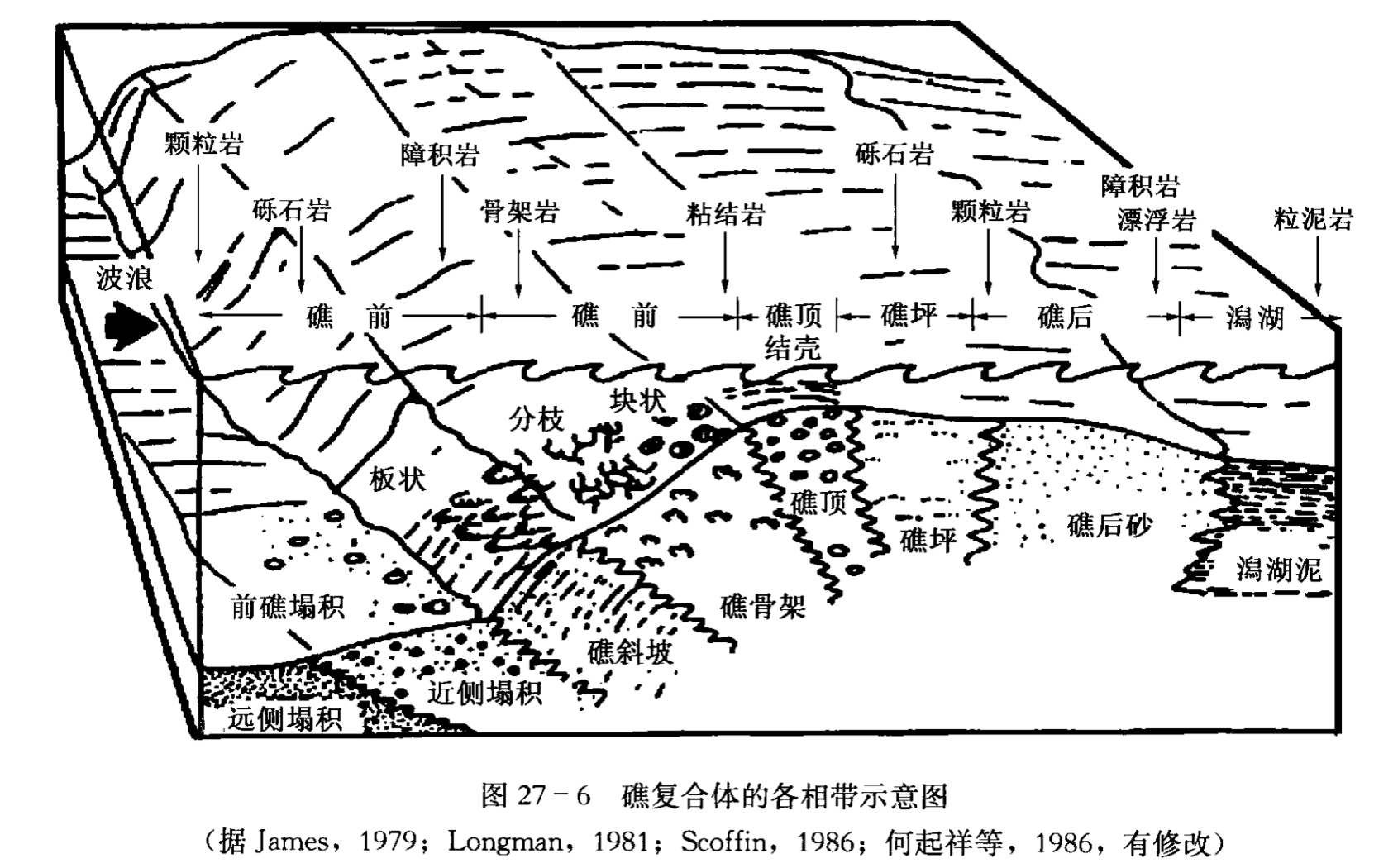
表27–2 现代礁复合体相的特征 (据何起样等,1986,有修改)
| 相 | 沉积作用和生物的控制 | 保存的生物类型 | 颗粒大小 | 分选性 | 骨架含量,% | 深度, m | 主要岩石类型 |
|---|---|---|---|---|---|---|---|
| 潟湖 | 低能, 生物浅穴发育,零星的流水和扰动 | 软体动物、棘皮、小粟虫、有孔虫和介形虫 | 泥,但混有粗骨架碎屑 | 差 | 0 | 5~30 | 颗粒泥质岩 |
| 礁后砂 | 有零星的风暴和流水横过礁, 含盐性, 重力滑动 | 仙掌藻、小粟虫、少量红藻和指状珊瑚 | 粗 | 中一好 | 0 | 1~10 | 颗粒岩 |
| 礁坪 | 零星风暴, 水循环好,分离出现 | 指状珊瑚、红藻、绿藻、大的底栖有孔虫 和大的珊瑚 | 粗—很粗 | 中 | 0~10 | 1~3 | 颗粒岩、有少量珊瑚 |
| 礁顶 | 波浪能量低, 持续扰动, 水循环好 | 抗浪的珊瑚和藻 | 很粗 | 中—好 | 0~80 | 0~2 | 颗粒岩 (少量粘结岩) |
| 礁骨架 | 水循环好, 波浪 能量强 | 丰富的珊瑚、藻、软体动物、棘皮、有孔虫 | 骨架和砂 | 差, 洞穴中有泥 | 28~80 | 1~30 | 粘结岩 |
| 礁斜坡 | 光线受限制, 有零星扰动, 碎屑的重力搬 运作用 | 软珊瑚、扁平状板状珊瑚, 海绵 | 混合的 | 差 | 5~40 | 20~50 | 泥 质 颗 粒 岩、粘结岩 |
| 近侧塌积岩 | 零星扰动, 重力搬运,缺少光线, 不稳定——稳定 | 仅有少量的活着的生物 | 中一粗 | 差—好 | 0 | 40~100 | 颗粒岩、泥质颗粒岩 |
| 远侧塌积岩 | 静水, 无光线, 沉积物重力滑动 | 浮游有孔虫 | 细 | 中—好 | 0 | 100~200 | 泥质颗粒岩 |
各相带的沉积特征分述于下。
一、礁骨架相
礁骨架相位于礁的前缘,是波浪和水流强烈扰动的环境,沉积水深多浅于30m(图27-6)。温暖的气候、富含营养的水流、充足的阳光以及水的循环条件,都适合于骨架生物生长,是造礁生物繁衍最旺盛的地方。如果拓殖的生物是钙质生物,那么生物的大量繁殖就可以造成生物骨架的发展和礁复合体的形成。但是,由于生物的侵蚀作用以及波浪、潮汐和流水的作用,常常使这些碳酸盐骨架遭到一定程度的破坏。一些破碎的原生骨架碎屑可从礁骨架搬运到礁后区去,或者在重力作用下堆积在礁前。这样,当骨架生长时,一方面有原生骨架保存下来,另一方面骨架间和骨架内的原生孔隙中也可以堆积大量的骨骼碎屑。一些研究者认为,在成熟的礁中,这种礁骨架碎屑总是在体积上比骨架自身的体积要大得多 ( Ladd,1950、1971)。
从现代礁的观察可以知道,在礁骨架相中有50%~100%的地方被骨架所覆盖。但是,当这些广泛延伸的原生骨架形成至埋藏后这段时间内,它们要受到不同程度的物理作用 (波浪、潮汐和水流)、生物作用(主要是钻孔) 以及成岩作用 (主要是泥晶化作用) 的影响,使这些原生骨架遭受不同程度的破坏。所以,礁骨架一般只有30%被保存下来 ( Longman,1981), 还有一些研究者认为连10%都不到 ( Land 和 Moore, 1977; Freidman, 1978)。
海南岛和西沙群岛的珊瑚礁也有类似的现象,原生骨架由于受波浪、潮汐以及生物钻孔作用而受到不同程度的破坏。
二、礁顶相
礁顶相是礁复合体中沉积水体最浅的相带 (0~2m),通常出现在礁的顶部。它可存在两个完全不同的沉积类型 (图27-6):
(1) 礁顶是由活着的珊瑚骨架组成,为扁平的板状珊瑚,低能区则由指状珊瑚组成。珊瑚呈扁平状可能是生存竞争的结果。因为在礁顶接近水面时,水体比较浅,只有呈扁平形状的珊瑚才能有效地利用一切空间,不至于在低潮时为周期性暴露在水面的恶劣环境下所扼杀。这种类型的礁可见于印度尼西亚和菲律宾。
(2) 由珊瑚砾块和红藻石组成。由于周期性暴露将杀死活着的珊瑚,故礁顶变成一个次生礁脊。珊瑚碎屑的粒度为砂到巨大的漂砾,漂砾是由风暴从礁骨架上撕裂下来的。停滞在礁顶上的珊瑚砾块通常要受到生物的钻孔,与此同时,还要受到藻、蠕虫、有孔虫以及其他生物的结壳。许多生物可以长在珊瑚的砾块下或砾块之间,这些生物的分异度比较差,活着的珊瑚非常少。然而海星、某些有孔虫以及藻类仍然比较繁盛。
在西沙群岛,一般礁复合体无活着的礁顶存在 (何起祥等,1986),实际上都是一些次生礁顶,它与太平洋中大量发育的次生礁顶类似。
三、礁坪相
礁坪相是礁复合体中最宽的一个相带,沉积物较粗,沉积水体较浅 (0~3m) (图27-6)。该地区地形平坦,在特大低潮时,部分地区可以露出水面。礁坪相出现在礁顶相和礁后砂相之间的地带,该环境的特征是除了零星散布的块状珊瑚和指状珊瑚外,更多的是珊瑚碎片以及分散的海草。与礁骨架相和礁顶相相比,礁坪相的波浪和水流的能量较低,缺少底质,水的循环受到限制,这就在一定程度上限制了生物的分异度和那些营滤食方式生活的生物的繁殖。但局部地区珊瑚较丰富,可以形成斑礁。此带阳光充足,促进了一些生物的生长。这些生物包括绿藻 (仙掌藻)、分枝状和节片状的红藻以及许多非钙质藻。海藻和海藻层的发育为各种有孔虫、潜穴生物,特别是软体动物提供了一个良好的栖息场所。海参、甲壳类、鱼以及啃食动物也很普遍。
礁坪宽度的变化取决于先前的地貌、礁体发育的时间、礁前缘的波能、陆源物质的注入量以及其他因素。伯利兹附近的斑礁的礁坪只有几米宽,而世界其他一些礁的礁坪宽度可达100m以上。西沙群岛的礁坪宽度一般都在100m以上,最宽的可达1~2km(包括一部分礁后砂相)。
礁坪上的沉积物分选中等,颗粒呈棱角状到次圆状。它们多由珊瑚碎屑、红藻、软体动物、棘皮类以及有孔虫组成。绝大部分的泥已被冲走,但是,球粒可以和海草一起共生。礁坪上有时也可以有大的珊瑚岩块和生长状态的块状珊瑚,通常它们都有被生物钻孔的痕迹。
四、礁后砂相
礁后砂相位于礁坪后侧,两者是逐渐过渡的 (图27–6)。波浪横过礁坪,其能量将大为降低。由于沉积物变细和不稳定,以致于使固着的营滤食方式生活的珊瑚不能正常繁殖。该处水深一般为1~5m,最深可达10m,有时其部分地区可以暴露在水面之上成为小岛。间歇性的风暴能把礁骨架破碎的物质从礁复合体的向海地带搬运到礁后环境中,同该处生长的生物,如软体动物、藻(主要是仙掌藻) 以及有孔虫等混在一起,沉积物分选中等到较好。现代珊瑚礁内,礁后砂的主要成分是珊瑚和钙藻碎片,但也常常见到棘皮类、软体动物和有孔虫的碎片,灰泥很少。
此相带宽度一般为数十米,但当海平面长期稳定时,可以造成几公里宽的礁后砂坪。在这些砂质礁岛中,通常有淡水透镜体,它们对碳酸盐砂的成岩变化起着重要的作用。
五、潟湖相
潟湖是指环礁内或礁复合体之后一个静水环境,其沉积物以灰泥为主 (图27–6)。潟湖相可以是礁复合体的一部分,也可不是礁复合体的一部分,这视其沉积物的来源是否与礁复合体有关。在西沙群岛,潟湖沉积主要是砂屑和泥屑。潟湖内还有少量的斑礁。潟湖的周围藻席比较发育。这些潟湖沉积物均来自礁体,故西沙群岛的潟湖也属于礁复合体。
潟湖的水深只有几米到几十米,波浪的能量比较低,水的循环受到限制,沉积物一般为碳酸盐泥和细粒的碳酸盐砂,分选差。除了来自礁骨架的极细粒的生物碎屑外,主要的生物碎屑是软体动物、有孔虫以及仙掌藻,缺少广海的生物。还有许多生物进行着广泛的掘穴活动,如棘皮类、甲壳类以及软体动物。
另外,还有一些潟湖,例如伯利兹堡礁南部的一些潟湖,它们具有深水陆架泥的特征,并含有浮游有孔虫 ( Scholl和 Kling,1972),这就和礁复合体成因的潟湖有所不同了。
六、礁斜坡相
礁斜坡相处于成熟或未成熟的礁复合体的礁骨架相向海一边,其特征是有一个较陡的斜坡 (图27–6)。在斜坡上,只有零星的珊瑚,当有海水扰动时,可发育八射珊瑚。水深通常有几十米 (表27-2),该处保持一个相对波浪能量比较低、阳光不太充足的环境。在这样一些因素的影响下,最适合于快速生长的软珊瑚的发育,而不利于石珊瑚的生长,尽管在该环境中石珊瑚并不一定缺失。为了获取有限的阳光,这些珊瑚采取平板状形态生长,珊瑚虫长在平板的上面。
在一些斜坡相中,硬海绵是很重要的,特别是在有礁壁存在的地方。
礁斜坡相的沉积物主要来自礁复合体的浅水部分,它们通过重力作用,漂移和沉降进入到该环境中。沉积物的分选性中等到差,因为礁斜坡相的沉积物来自该相的下部和上部,粗细碎屑混合在一起。这些碎屑物质都遭受了不同程度的磨蚀作用和生物的钻孔作用。
西沙群岛礁复合体的斜坡相,斜坡的坡度较大,有时甚至直立,坡面坎坷不平,有时可见到冲刷、溶蚀构造。风暴破坏礁骨架带来的碎屑物质一部分被抛到礁坪上,另一部分顺着礁斜坡滑落,细小的碎屑被带到斜坡的下部。
通常礁斜坡相都有一个较陡的斜坡,其倾角一般为50°~90°。该相中的生物主要是仙掌藻、Zooantharians、海绵以及硬海绵,这些生物死亡后和沉积物一起堆积下来。斜坡沉积物的另一个特征是呈透镜状,下部比上部更厚。
要在古代岩石中鉴别礁的斜坡相比较困难。看来,珊瑚的板状形态可能是一个很好的标志,另一个标志是其近侧出现塌积岩。
七、近侧塌积岩相
近侧塌积岩的环境是指礁斜坡之下的那个地带,其特征是含有大量的、来自礁复合体的碎屑和少量活着的钙质生物 (图27-6)。水深变化很大,一般不能用潜水的方式进行研究。通常一些研究者使用潜水艇在该相带进行观察 ( Land和 Moore,1977)。由于它的深度比较大,所以在该环境中波浪能量低,光线微弱,甚至有时透不进光线。其沉积物主要是通过重力作用沉积的。一般情况下水体处于静止状态,只有沉积物降落时,才产生局部的流动。
礁的塌积物来自礁复合体。礁岩碎块有时很大,其粒径可达几米以上。此外还有一些仙掌藻、红藻以及其他各种不同的生物成因的颗粒。但是,在特殊情况下也有这样一些塌积沉积物,它们的碎屑是来自礁后潟湖和深水中任何一种主要的生物组分。岩石类型从泥岩到颗粒岩都有,但以骨骼泥质颗粒岩为主。在一些地区,一些陆源物质也可以混入到塌积岩中。
横向上连续性比较好的层状构造和零星的生物潜穴是塌积相中常见的沉积构造。层的厚度从几厘米到几米,通常被页岩所分隔。塌积岩中这种层状构造的存在是区别礁骨架相和礁后相的重要标志。
八、远侧塌积岩相
远侧塌积岩相位于塌积岩相的下斜坡 (图27-6)。该地区的沉积物粒度较细,含大量的浮游生物。它是浮游生物和来自礁复合体的细粒碎屑物质的混合沉积体。远侧塌积岩和近侧塌积岩之间是渐变的。当来自礁复合体的碎屑物质逐渐消失后,远侧塌积岩的塌积物就逐渐过渡为深水盆地相的沉积物。
表27-2综合了上述各相的沉积特征。
第三节 礁的分布规律及礁与油气关系
一、礁的分布控制因素
礁在海域和湖域中分布很广泛,但其都是在一定地质背景条件下主要由生物作用形成的地质体,因此其分布具有一定的规律。按生物礁的发生、发展、消亡以及最终形态和内部构造受不同条件控制的特点,主要从以下5个方面考察礁的控制因素。
(一) 海平面的升降
造礁生物生存在一定的海水深度,既不能露出水面,也不能生存于较深的水中。因此,组成礁的藻和一些植物,只能在光合带繁殖。当海平面保持稳定,礁就会沿自身的碎石斜坡侧向向海推进。如果海平面缓慢地上升,则礁基本上向上生长,而不会横向移动,或者向海岸方向逐渐推进; 如果海平面快速上升,礁则因水的深度迅速增大,无生存能力而消亡。如果海平面缓慢地下降,就会造成礁向海和向下移动; 如果海平面快速大幅度下降,就会造成礁死亡。
(二) 大地构造位置
相对稳定的构造背景有利于礁的生长发育。大地构造位置和构造活动强度对礁的控制意义与海平面的变化同等重要。礁的形成基本上是受构造大陆架的控制,这里的沉积作用是在浅水、无陆源碎屑的环境中进行的。在这个广阔的领域里,可以识别出4个主要亚类。第一种礁形成堡礁 (如西得克萨斯州的三叠纪礁) 或不连续的点礁。有时大陆架的边缘可能是断层,礁在断层悬崖的顶峰上建造,如英国北部克拉科和克拉文礁。第二种礁,因大陆架太深,使礁沿着背斜顶部同沉积移动发育而形成。利比亚的古新世因泰萨尔礁就是一个例子,英国的早石炭世博伍伦德槽的克里锡罗礁也是一例。第三种礁为海底火山喷发时在熔岩锥上形成,墨西哥塔菲拉勒盆地的泥盆纪生尔格达德是典型代表之一 ( Massa,1965)。第四种礁是大面积无规律分布的点礁,如加拿大地盾边缘上的志留纪点礁 ( Lowenstam, 1950)。
局部构造也同样影响礁的生长,例如海底的突起,在适宜条件下就可以成礁。特别是大陆架脊,更有利于礁的生长,例如墨西哥湾坎佩切滩即是发育于大陆架脊上。
断层也能控制礁的发育,断层的发生和发育可使礁伴随着发生位移。
(三) 地貌的影响
岸礁和环礁严格地受海底地貌和海岸带地貌特点的控制,平缓的地貌有利于生物礁的生长,例如我国南海的岸礁,其形态和分布特点均受海岸带坡度和海岸地形的制约。海南岛的珊瑚岸礁,根据地貌可分为平直海岸岸礁,弯曲海岸岸礁等。又如海台地、海山控制着太平洋的岸礁、堤礁的形态和分布。
(四) 沉积环境的影响
礁主要分布于南、北纬32°之间,少量的珊瑚藻礁分布于北纬49°。由此可见,礁的形成需要温暖的条件。除此之外,生物生长需要清洁的水域,以免有碎屑物质使生物窒息; 还需要有机物质和CaCO₃的供给,保证生物的造架作用。各种不同的生物生存的环境各有不同,例如珊瑚礁生长于水下,而蛇螺礁则主要生长在潮间带。不同的生物需要的盐度也各有不同,如珊瑚生长的海水盐度为20~40mg/L,红藻可在广泛盐度变化的环境中 (18~54mg/L) 生存; 牡蛎则生存于微咸水环境中 (10mg/L), 在河口中最发育。
不同类型的生物礁分布于不同的沉积环境,以最常见的岸礁、点礁、堡礁、宝塔礁、丘礁为例,其中岸礁分布于各种海岸带,点礁常分布于堡礁之后的潟湖中,堡礁分布于大陆架边缘地带,宝塔礁和丘礁分布于斜坡地带。
(五) 生物群落
生物群落也是控制礁分布的因素之一,而且是成礁的关键性要素。造礁生物能分泌大而茁壮的各种生态骨骼 (分枝状、半球状或板状),这些具有骨骼的生物在显生宙期间具有一定的分布规律。有些生物群落虽不具有骨骼,但它可以分泌粘液而形成丘礁。所以礁在各个时代都有分布,但组成的生物群落不同。
(1) 震旦纪:蓝绿藻。(2) 寒武—奥陶纪:绿藻、蓝绿藻及古杯动物、苔藓虫。(3) 志留纪:层孔虫、四射珊瑚。(4) 泥盆纪:四射珊瑚、管状珊瑚、层孔虫、苔藓虫。(5) 石炭纪:四射珊瑚、板状珊瑚。(6) 二叠纪:苔藓虫、水螅、海绵、珊瑚。(7) 三叠纪:六射珊瑚、钙藻、海绵、树枝状珊瑚。(8) 侏罗纪:六射珊瑚及藻类。 (9) 白垩纪:钙藻、珊瑚、苔藓虫、厚壳蛤、有孔虫。(10) 古近纪:珊瑚及藻。(11) 新近纪:藻、珊瑚、唇口目、苔藓虫。
二、中国的生物礁分布
我国地质学家对生物礁的研究表明,各地质历史时期均有生物礁发育。
(一) 震旦纪
我国分布广泛的震旦纪地层,包括南方的灯影组、北方的雾迷山组、塔里木盆地的奇格布拉格组都有藻礁发育,或称为点礁和藻滩。这种礁由藻类组成,规模较小,一般几平方米或几十平方米,根据叠层石和几何形态很容易辨别。
(二) 寒武纪
已在扬子地台北缘和湘西渔塘早寒武世地层中发现礁,前者为补丁礁,后者为堤礁。补丁礁(岸礁) 基本上由网格古杯 ( Retecyathus)组成,规模较小,厚仅几米,叠覆成复合体。堤礁主要是由灌木丛状表附藻和管状葛万藻以及间接参与造礁的各种蓝绿藻组成,附礁生物仅见少量三叶虫和介形虫。礁体宽0.2~0.8km,长4~6km,厚几十米。
(三) 奥陶纪
在下扬子区 (包括钱塘区) 和塔里木盆地已发现多处生物礁,主要由灌木丛状、管状钙藻类 (可能属红藻) 和苔藓虫、珊瑚、层孔虫等造礁生物组成。礁体厚几十米,延伸最长达11km, 属于堤礁性质。
(四) 志留纪
志留纪生物礁见于扬子地台北缘的陕西宁强、四川广元、城口,为点礁。主要由蜂巢珊瑚组成,也有层孔虫和苔藓虫,伴有丰富海百合和其他生物。此外,在贵州黔中古隆起周围也有志留纪点礁分布,厚达50m左右。
(五) 泥盆纪
泥盆纪是我国南方古生代生物礁最发育的时期,在贵州、广西、四川、湖南等地均有发现。以贵州为例,生物礁主要分布于台地边缘相带,造礁生物主要为层孔虫和珊瑚,附礁生物有棘皮、苔藓虫、腕足、红藻等,生物含量达20%~90%。厚达240m,延伸几公里。可十分清楚地划分出礁核相、前礁相、后礁相。广西大厂龙头山泥盆纪生物礁的造礁生物与贵州的基本一致,但其发育于盆地的隆起部位。
生物层礁,高几米至数十米,宽几公里至数十公里。主要造礁生物为层孔虫和珊瑚。
台地边缘礁,高达百余米。造礁生物主要是六方珊瑚和板状层孔虫,附礁生物有海百合与腕足等。生物含量达60%~80%。
丘状礁,发育在台地海盆相带内,形成于隆起之上。造礁生物以珊瑚、层孔虫为主。
(六) 石炭纪
已在广西地区大塘期中发现点礁 (方少仙等,1985)。主要造架生物是四射珊瑚与苔藓虫,附礁生物是海百合、腕足、有孔虫、介形虫、绿藻。礁核厚达 114m,分布直径达200m。发育于孤立的碳酸盐台地上。
(七) 二叠纪
二叠纪是我国生物礁比较发育的时期,特别是我国南方,已在云南、湖南、四川、湖北等地区发现二叠纪的生物礁,分布广泛,类型多样,生物丰富。礁的类型有:(1) 台地边缘生物礁,如云南省广南堇那孟生物礁。造礁生物主要是钙质海绵,附礁生物为有孔虫、腹足类、腕足类、苔藓虫等。川东、鄂西二叠纪生物礁也属此类。(2) 点礁,如滇黔桂南盘江地区的孤立碳酸盐台地上的生物礁。造礁生物以海绵为主,苔藓虫、水螅、珊瑚为次。又如川东一带,不少礁属于台地内的点礁。
(八) 三叠纪
已在贵州和四川西北部等地发现有三叠纪生物礁。贵州三叠纪生物礁属堤礁 (堡礁),形成于安尼锡克期和拉丁尼克期,造礁生物以红藻为主,附礁生物有腹足类、腕足类、有孔虫等,厚度达几百米。川西北龙门山前晚三叠世的生物礁属于点礁 (补丁礁),造礁生物以海绵为主,附礁生物有腕足、棘皮、介形虫等。呈礁群分布,礁残留高度约25m,椭圆形,出露宽度约50m, 礁间距100~500m不等。
(九) 古近—新近纪
珠江口盆地古近一新近纪发育有大量的海相生物礁群,属于塔礁、点礁,推测尚有台地边缘礁。造礁生物主要有红藻,珊瑚、海绵、苔藓虫及水螅,其中最主要的是红藻。附礁生物有有孔虫、腕足、软体,棘皮、介形虫等,厚达百余米,生物礁群面积近千平方公里,有的已经钻探证实为油田。除此之外,莺歌海盆地和北部湾盆地均发育有新近纪生物礁。
三、礁分布与油气的关系
礁及其复合体极易形成有效圈闭而成藏,可以说凡有碳酸盐发育的地区,大部分都存在由礁控制的油气田,因此可见礁与油气关系密切,且礁具有独特的富油气特征。
(一) 礁常是成群或成带分布
墨西哥埃尔阿布拉环礁宽近80km,长180km,现今陆上分布在黄金巷带上的油气田约有50个,海上已发现了20个油气田。又如美国西得克萨斯马蹄形礁,位于米德兰得内克拉通盆地北端,地下延伸282km,面积约15540km²,是世界上最大的礁群之一。沿环礁顶部已发现有15个油田,可采储量达 3.5×10⁸t。美国密执安盆地中的中、晚志留世,在盆地边缘发育有堡礁,大陆架滨外形成宝塔礁。每个宝塔礁平均为0.5km²,其高达90~180m,围绕密执安盆地分布达数千个 ( Shaver,1977)。这些宝塔礁聚集有大量的油气,加拿大阿尔帕达盆地雨虹油田也是由许多中泥盆世的礁群组成。
礁群或礁带不仅在平面上有这样分布的特点,而且在垂向上也成群体分布。众所周知,单个礁体有发生、发展和消亡(定殖、拓殖、泛殖、统殖) 的过程。这样的过程受着环境和亚环境的控制,从而礁体随着环境和亚环境的不断变化而不断形成,所以在垂向上也形成多个礁体的复合体。
(二) 礁储层常具有较高的孔隙度和渗透率
碳酸盐岩储层通常具有较低的孔隙度和渗透率。礁型储集体相反,常具有异常高的孔隙度和渗透率,比良好的砂岩储层的孔隙度和渗透率还高。
世界上有8口日产万吨的油井,其中有4口是产自礁型油田 (墨西哥黄金巷3口,利比亚伊特里斯1口),这些油田具有很高的孔隙度和渗透率的礁储集体是重要的因素。
礁型油气田一般都具有大于10%的孔隙度,渗透率在100x10-3 um2以上 (表27-3)。因此礁型油气田单井产量常常很高。这是由于礁体白云岩结构构造的特殊性所致。
表27-3礁型油气田物性特征
| 国 家 | 油 田 | 孔隙度,% | 渗透率, 10⁻³μm² |
|---|---|---|---|
| 伊拉克 | 基尔库克 | 7~25 | |
| 利比亚 | 英蒂萨斯 | 22 | 4~500 |
| 美国 | 利史纳得 | 7.6 | 19.4 |
| 加拿大 | 邦尼格仑 | 9.55 | 115~1271 |
| 加拿大 | 列杜克D | 8 | 100~1000 |
| 加拿大 | 金穗 | 15 | |
| 加拿大 | 朱迪湾 | 12.5 | 170 |
| 加拿大 | 虹G | 10.6 | 565 |
| 中国 | 建南 | 14.6 | |
| 中国 | 滨南 | 46 (礁核) |
对整个礁体而言,孔隙度和渗透性相当不均。礁核相带孔隙度和渗透性最高,而礁侧相(后礁相、前礁相) 孔隙度和渗透率则很低。如伊拉克基尔库克油田,后礁相--潟湖相灰岩岩性比较致密,物性很差; 礁核灰岩孔隙度、渗透率很高; 前礁灰岩由于白云岩化作用,使孔隙度、渗透率增高。利比亚英蒂萨斯礁块油田,礁块的中央 (礁核部分) 孔隙度高达22%~26%,而礁侧相孔隙度则下降到15%。又如我国东营凹陷滨南陆相碳酸盐岩礁型油田,礁核孔隙度可达46%,而礁侧相为6%左右。世界许多礁型油田都具有类似的特点。
值得注意的是,在实际中有不少礁型油气田,由于白云岩化作用和充填胶结作用,使礁块的物性特点发生异常。
(三) 礁复合体发育良好的生、储、盖组合
礁块的存在是沉积环境的一个标志。在向岸一方常常是潟湖沉积环境,从礁向广海 (广湖) 一侧则是盆地沉积环境。
许多资料表明,前礁的盆地相和后礁的潟湖相都是有机物质丰富的细粒碳酸盐沉积。这些富含有机物质的石灰岩或泥灰岩,可以在有利的条件下形成烃源岩。有的礁体直接就为黑色页岩所围,这些黑色页岩可能是良好的烃源岩。因此,礁的周围常常是生油岩系。不论是海进或是海退沉积序列,在礁块上方可上覆蒸发岩系或细粒碳酸盐岩、页岩,形成良好的盖层。
综上所述,礁型油气田具有良好的生、储、盖组合条件。在有利条件下,必然形成良好的礁型圈闭和油气田。
(四) 礁核常构成礁型油气田
这是由于礁核相具有最好的孔隙度和渗透性,所以在勘探礁型油气田时,一定要搞清礁体相带的分布。但是经常也可以看到油气的分布不是受礁核相带的制约,而是受白云岩化作用的控制。例如我国建南礁型气田,生物礁发育于上二叠统长兴组中部。礁高156m,面积约15 km2。造礁生物以海绵、层孔虫、蓝绿藻为主。储集体的储集空间是以次生白云岩的晶间孔隙和溶蚀孔隙为主,孔隙度达14.6%,而生物骨架灰岩则具有很低的孔隙度。因此,建南气田的天然气分布受白云岩化作用的制约。
应注意油、气、水在礁圈闭中的差异聚集,例如美国密执安盆地宝塔礁带的油、气、水的分布,向盆地方向分布的礁体储存天然气,向盆地边缘则依次为油、水。这是由于流体在礁群中向上倾方向运移分异捕获所致,这些礁群型圈闭为同一动力学系统,通过下伏地层水动力连接起来 (Gill, 1979)。
总之,礁及礁复合体的分布是控制油气分布及油气成藏的最重要的条件之一,也是特殊的礁圈闭形成的基础,它的发育演化和空间分布规律为油气聚集提供了良好的圈闭条件。