1 䗴类有孔虫
内容节选自《古生物学》(童金南主编,2021年版)第八章 微体古生物
䗴类(fusulines)又名纺锤虫,是晚古生代低纬度浅海区十分繁荣、具有特殊复杂壳体结构的一类单细胞原生动物,分类位置属于有孔虫纲䗴目。䗴类在地质时代仅分布于石炭纪至二叠纪,其演化迅速,很多是标准化石,有重要的生物地层学意义。䗴类生活于水深100m左右的热带、亚热带平静正常的浅海环境,为浅海底栖生物。
䗴壳的形态及构造
䗴类为多房室包旋式壳(involute test,即外层壳圈全部包裹内层壳圈,最后在壳外只能看到最外面的壳圈),圆形的初房位于壳中央。初房上的圆形开口是细胞质溢出的通道。细胞质不断增长并阶段性地分泌壳质形成的外侧壳壁叫旋壁(spirotheca);旋壁围绕一条假想轴增长,同时向轴的两端伸展,包裹初房及内圈的房室,此轴称为旋轴(axis)。旋壁前端阶段性地向内弯折形成前壁(antetheca),前壁基部的向外开口为口孔(aperture)。如果只有一个口孔,则称为单口孔(single aperture);如果有多个口孔,则称为复口孔(multiple apertures)。虫体生长一段时间后,细胞原生质会从口孔流出,贴附在前壁前方分泌新的外壁和新的前壁;与此同时,先前的前壁就变为隔壁(septum),而该隔壁与新前壁之间的窄长空间就成为新的房室(chamber)。䗴类按此方式依次增长形成多房室。旋壁绕旋轴一圈即构成一个壳圈(whorl)(图8-5之1)。
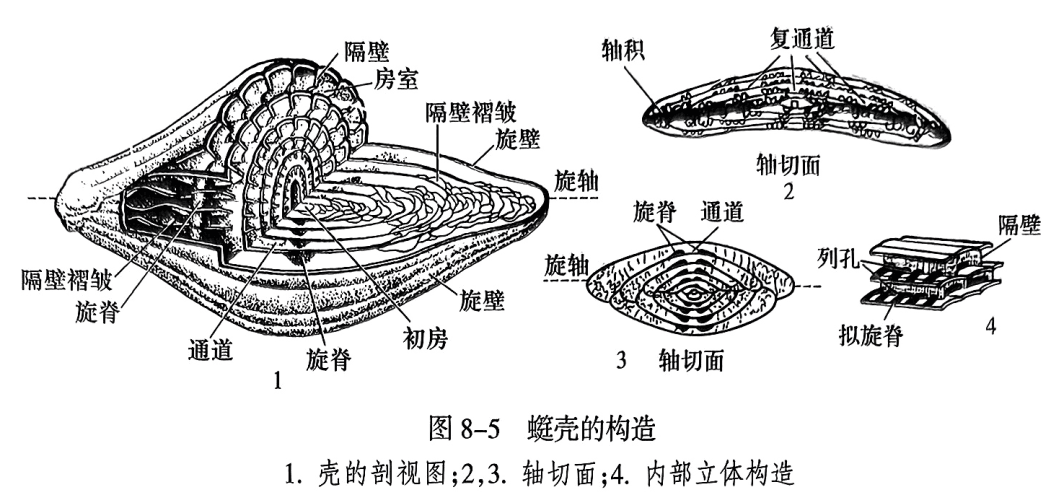
当前壁被新生的房室包裹转变为隔壁后,先前向外开口的口孔就变为隔壁基部的隔壁孔(septal pore)。先后形成的隔壁孔从内向外彼此贯通就形成一条单通道(tunnel),或多条复通道(multiple tunnels);通道是细胞原生质流动的通道,其两侧通常有次生堆积物,以限制原生质的流通;这些次生堆积物呈隆起的脊状随通道从内到外盘旋生长,在只有单个通道的䗴壳中在通道两侧形成两条成对的隆脊,称为旋脊(chomata)(图8-5之1~3);而在具有复通道的䗴壳中就形成了多条隆脊,称为拟旋脊(parachomata)(图8-5之4)。部分䗴类壳体中沿轴部可有次生钙质物充填,叫轴积(axial filling)(图8-5之2)。
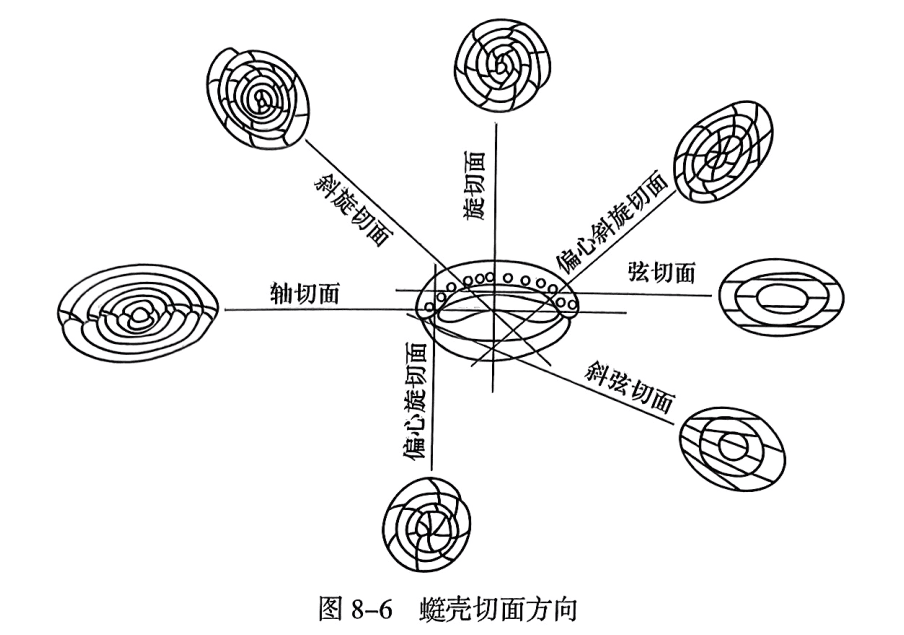
由于䗴类为包旋式壳,对其内部构造的观察和研究需要做不同位置和方向的切面。一般采用下面三种切面(图8-6):①轴切面(axial section),通过初房并平行于旋轴的切面;②旋切面(spiral section),通过初房并垂直于旋轴的切面,也称为中切面;③弦切面(tangential section),未通过初房但平行于旋轴的切面。显然,在各种切面上能够观察到的䗴壳构造是不同的,例如,在轴切面和旋切面上都可以观察到所有的壳圈,包括壳圈的数量,壳圈生长的均匀性,以及内外壳圈包裹的紧密程度等。但在轴切面上只能看到旋向生长的构造,而在旋切面上只能观察轴向排列的构造。弦切面虽然与轴切面一样可以观察旋向生长构造,但它无法知道整个䗴壳到底有多大和一共有多少个壳圈。因此,要完整地知道整个䗴壳的内部构造特征,一般需要借助多个方向的切面综合观察。当然,在实际工作中,还有大量的切面是偏离以上三种类型斜切面(obliquesection)的,在这种情况下,通常要尽量寻求与轴切面最为接近的切面。
近年来,由于CT扫描技术在化石研究中的应用,人们可以通过 Micro-CT对䗴壳扫描,建立其内部的立体结构,从而更加全面地了解其构造全貌。不过,目前这种技术还未能在䗴类化石研究中得以广泛应用,其主要难题是:䗴类化石都是钙质壳体,其主要保存在碳酸盐岩地层中,壳体与围岩的密度差异不大,当前的CT扫描技术尚不能将两者有效地区分开。
䗴壳形态构造的变化
(1)壳形的变化
䗴壳形状多样,常呈纺锤形或椭圆形。人们以长、宽等参数表示它的形状。壳长指平行于轴向上壳的最大长度;壳宽是垂直于轴向上壳的最大宽度。在轴切面上可以准确地测定䗴壳的壳长和壳宽。壳长/壳宽,即轴率(axialratio)。按轴率可将䗴壳分为长轴型(即轴率>1,为纺锤形)、等轴型(轴率=1,为球形)、短轴型(轴率<1,为透镜形)三类。有少数䗴类晚期壳圈松开,呈喇叭形。
(2)旋壁的变化
䗴壳为钙质微粒状分层壳,旋壁由原生壁和次生壁组成(图8-7)。前者包括致密层、透明层及蜂巢层;后者包括内、外疏松层。
致密层(tectum),是薄而呈黑色的密度较大的层,在显微镜下呈一条黑线。所有的䗴壳都具有致密层。
透明层(diaphanotheca),在致密层之内,为一浅色透明的壳质层。
蜂巢层(keriotheca),位于致密层之内,为一较厚且具有蜂巢状构造的壳层,在垂直旋壁的切面上呈梳状。
疏松层(tectorium,复数为 tectoria),通常为不太致密和不均一的灰黑色层,附在其他壳层的表面。位于致密层之外的叫外疏松层(outer tectorium),位于壳壁内表面的叫内疏松层(inner tectorium)。终壳圈外表面不见外疏松层,说明它们是一种次生堆积物质。
不同䗴类具有不同的旋壁构造,归纳起来可分为以下几种类型(图8-7之7~11):
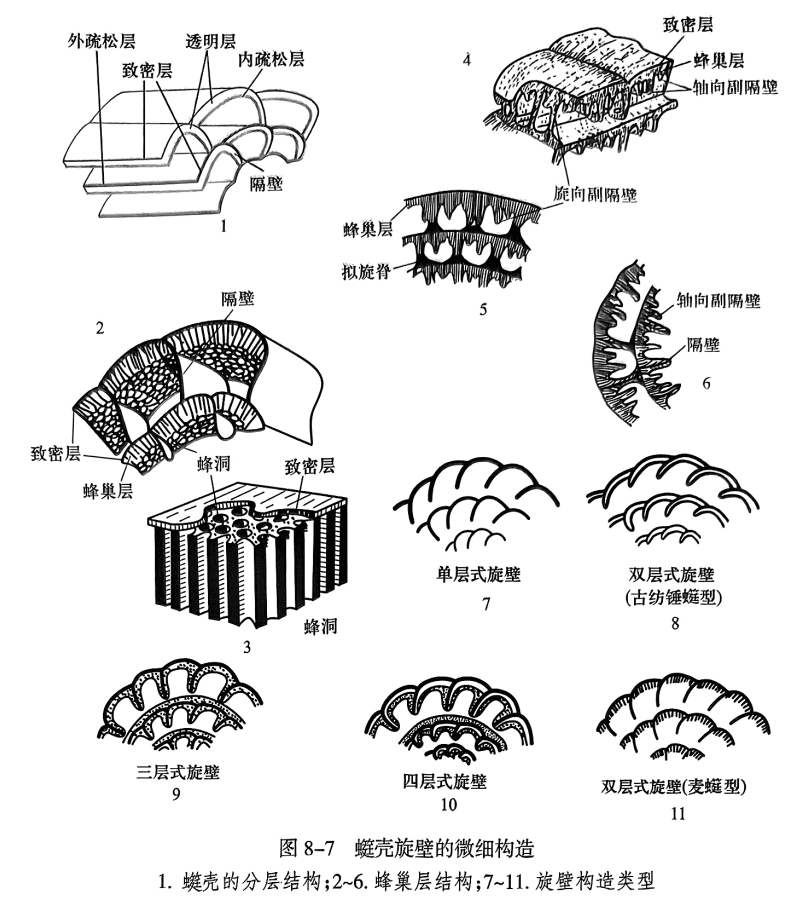
单层式(monolamellar),旋壁仅由一致密层组成。有些原始䗴类的旋壁仅由一层浅灰色的疏松物质组成,称为原始层(protheca)。
双层式(bilamellar),可分为两种类型:一是由致密层及透明层组成,称为古纺锤䗴型(palaeofusulinid)旋壁;二是由致密层及蜂巢层组成,称为麦䗴型(triticitid)旋壁。
三层式(trilamellar),也可分为两种类型,旋壁由致密层和内、外疏松层组成的称为原小纺锤䗴型(profusulinellid)旋壁。在一些高级䗴类中,旋壁由致密层、蜂巢层及内疏松层组成,称为费伯克䗴型(verbeekinid)旋壁。
四层式(quadrilamellar),旋壁由致密层、透明层及内、外疏松层组成,称小纺锤䗴型(fusulinellid)旋壁。
(3)隔壁的变化
䗴类隔壁可呈平直状或褶皱状。褶皱状的隔壁从基部和两端褶皱发展到全面褶皱,隔壁褶皱的强烈程度因属种而异。由于隔壁是轴向伸展的构造,因此没有褶皱的平直状隔壁在轴切面上是不能被观察到的。但隔壁一旦出现褶皱,就立即能够在轴切面上看到,因此轴切面(及弦切面)是判断隔壁褶皱程度的最重要的观察切面。
(4)副隔壁
在二叠纪出现的许多䗴类中,其蜂巢层局部规则地聚集下延形成比隔壁略短的薄板,称为副隔壁(septulum,复数 septula)。与旋轴平行的副隔壁叫轴向副隔壁(axialseptulum);与旋轴垂直的副隔壁称旋向副隔壁(spiral septulum)。如新希瓦格䗴(Neoschwagerina)具有两种副隔壁(图8-7之4~6)。
䗴类的演化趋向
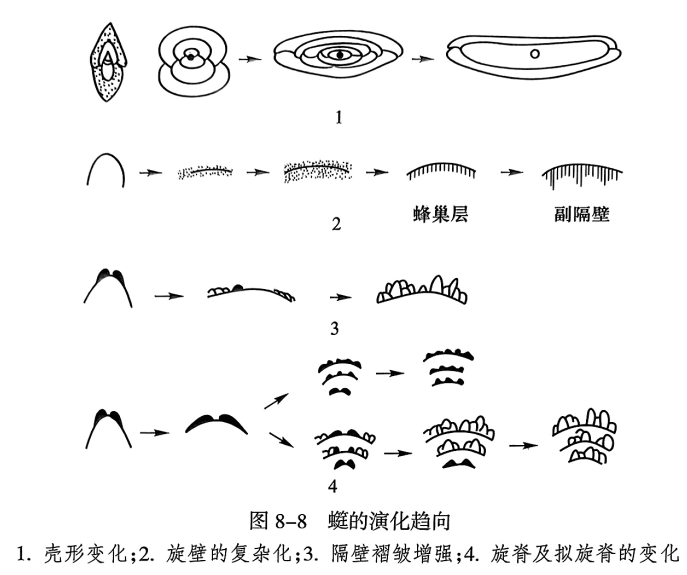
䗴类有孔虫在早石炭世晚期出现,至二叠纪末灭绝。在这段不太长的地质历程中,䗴壳构造却发生了迅速而有规律的变化,这为研究䗴类的演化提供了有利的条件。现将䗴壳的一些主要演化趋向归纳如下(图8-8)。
(1)壳体不断增长。低级䗴类的壳微小,壳的直径不到1mm,而绝大多数高度发展的䗴类都比较大,有的可比低级类型的体积大几十倍。在壳体由小变大的同时,䗴类的形状也由短轴型如凸镜形、盘形变为等轴或长轴型,如球形、纺锤形、圆柱形等。
(2)旋壁构造复杂化。旋壁构造的变化是䗴类演变中极为重要而明显的现象。原始䗴类的旋壁由未分化的单层式构成;以后的䗴类渐变为双层式、三层式,之后进一步发展成四层式。晚石炭世旋壁构造中蜂巢层的出现是䗴类演化中一个重要转变现象,它们起到加固壳壁的作用。早二叠世晚期副隔壁的出现又是䗴壳加固的另一种新形式,演变出新希瓦格䗴科的许多代表属种。
(3)隔壁和旋脊变化。这是䗴类演化的另一种形式。较原始的䗴类的隔壁平直且旋脊发育,随后出现两极褶皱,进而全部变为强烈褶皱;与此同时,旋脊也随之减弱,乃至消失,这是纺锤䗴科的演变特征。另一部分䗴类则在演变过程中隔壁不出现褶皱,但其进步类型发育出拟旋脊、副隔壁,这是费伯克䗴科的演变特征,说明它们与前者不是一个演化系列。
有孔虫的地史分布
有孔虫从前寒武纪末期出现直到现代,其总的演变趋势是逐步繁盛,在整个发展过程中形成了几个发展演化阶段,其中有三个重要的繁盛期,具体如下:
(1)寒武纪-泥盆纪是有孔虫的初步发育时期。寒武纪和奥陶纪的有孔虫个体微小,壳质脆弱,构造简单,全部为单房室壳类型,它们属于网足虫目及串珠虫目的一些原始类型。在志留纪有孔虫有了进一步发展,以胶结壳有孔虫为主。在泥盆纪胶结壳有孔虫又出现了不少新的种类,钙质微粒状壳有孔虫以内卷虫目为主。
(2)石炭纪-二叠纪是有孔虫的第一个繁盛时期。石炭纪有孔虫动物群的主要特点是䗴类的出现,并得到极其迅速的发展,具有十分重要的地层意义;胶结壳及钙质壳有孔虫大幅度发展,似瓷质壳有孔虫也在早石炭世出现。早一中二叠世䗴类发展到极盛,但到二叠纪末灭绝。同时,内卷虫目中除个别科的代表延到三叠纪外,其余全部也于二叠纪末灭绝。
(3)中生代的有孔虫以轮虫目占优势。具有钙质透明微孔壳的有孔虫,在三叠纪占主要地位。在侏罗纪时,有孔虫进化史上的重大事件是浮游有孔虫的出现。
(4)白垩纪是有孔虫的第二个繁盛时期。新生代的有孔虫类别大约有1/4在白垩纪已开始出现,其中大多诞生于晚白垩世。但也有许多仅生存于白垩纪的标准化石,尤其是浮游有孔虫成为划分对比白垩纪海相地层的重要依据。白垩纪有孔虫在数量上也是十分丰富的,是重要的造岩生物。
(5)古近纪是有孔虫第三个繁盛期。在大量有白垩纪特色的科属灭绝后,古近纪早期(古新世)又有一批新类型先驱种类开始出现。始新世是古近纪有孔虫的极盛时期,浮游有孔虫中出现了一些仅限于始新世的标准化石,同时底栖大个体有孔虫在许多地区成为造岩生物,形成货币虫灰岩、蜂巢虫灰岩等。底栖小个体有孔虫也出现了很多新类型。渐新世的有孔虫比始新世稍为逊色,原有的许多重要的浮游类型开始灭绝。新近纪的有孔虫基本上具备了现代有孔虫动物群的面貌。