8.1植物的形态结构
植物的形态、结构复杂。植物除原始类群外,都已分化出根、茎、叶和生殖器官等部分,并具有起输导作用的维管系统。
一、根
根(root)是植物的营养器官,它将植物固定在土壤中,并把从土壤中吸收的水分和无机盐输送到植物体的地上部分。根的形态除因类别而不同外,还常随环境而异。旱生植物的根系能扎入深层土壤或膨大;潮湿地区植物根系较浅,常水平延伸或在茎的下部形成不定根或板状根以加强支撑。根部化石最常见于煤层的底板层,在古环境分析中具有独到作用。
二、茎
茎(stem)是连接叶和根的轴状结构,一般生长在地面以上(也有些生于地下或水中)。茎的功能是输送水分、无机盐和有机养料,支持树冠。
茎的生长方向与根相反,为背地性,一般垂直向上生长。由于生态环境变化,茎的形态结构比较复杂。除绝大多数植物是直立茎外,在暖湿地区还常发育横卧于地面的匍匐茎和附着它物的攀援茎和缠绕茎。有些植物除地上茎外,还有埋在土中的地下茎。按茎的生活类型,植物可分为具有高大显著主干的乔木(arbor),主干不明显而较矮的灌木(frutex),攀附他物的藤本(liane)(木质和草质),矮小、无木质茎的草本植物(herbaceous plant),以及寄生其他植物体的附生植物(epiphyte)等。
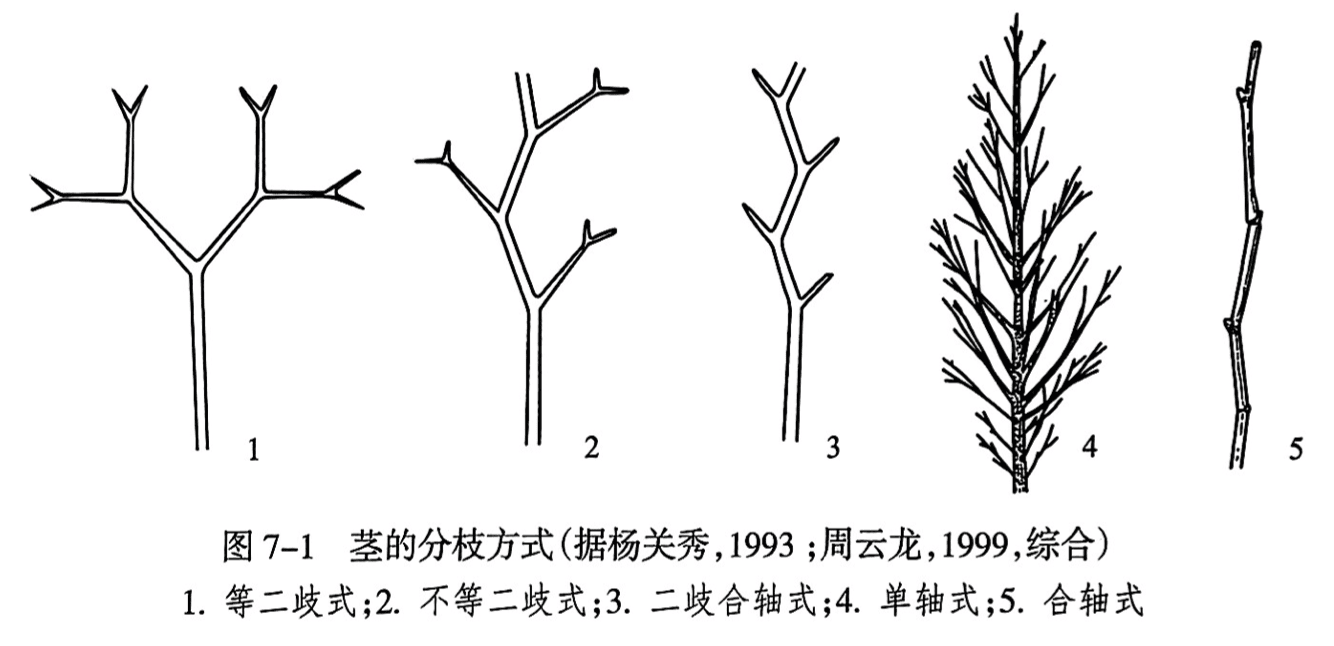
茎的基本分枝方式有二歧式和单轴式两大类型。二歧式分枝(dichotomus branching)是由茎顶端分生组织分生出两个大致相等的顶端发育而成,可进一步分为等二歧式分枝和不等二歧式分枝(图7-1之1.2)。等二歧式分枝不断重复形成茎干系统;但在相当多的情况下,两个顶端发育不相等,就形成不等二歧式分枝。明显的不等二歧式分枝就形成了“之”字形的轴和较短的“侧枝”,称为二歧合轴式分枝(dichopodial branching)(图7-1之3)。有的学者认为二歧合轴式分枝是植物演化历史上的重大事件,它导致单轴式分枝、羽状的叶子和羽状脉序的中脉形成。单轴式分枝(monopodial branching)有明显的主轴和由主轴分出的侧枝(图7-1之4),裸子植物和一些被子植物如杨树等为这种分枝方式。种子植物的合轴式分枝(sympodial braching)被认为由单轴式分枝进化而来(图7-1之5)。禾木科植物的分枝方式具有特殊性,其分枝集中发生在接近地面或地下的茎节上。
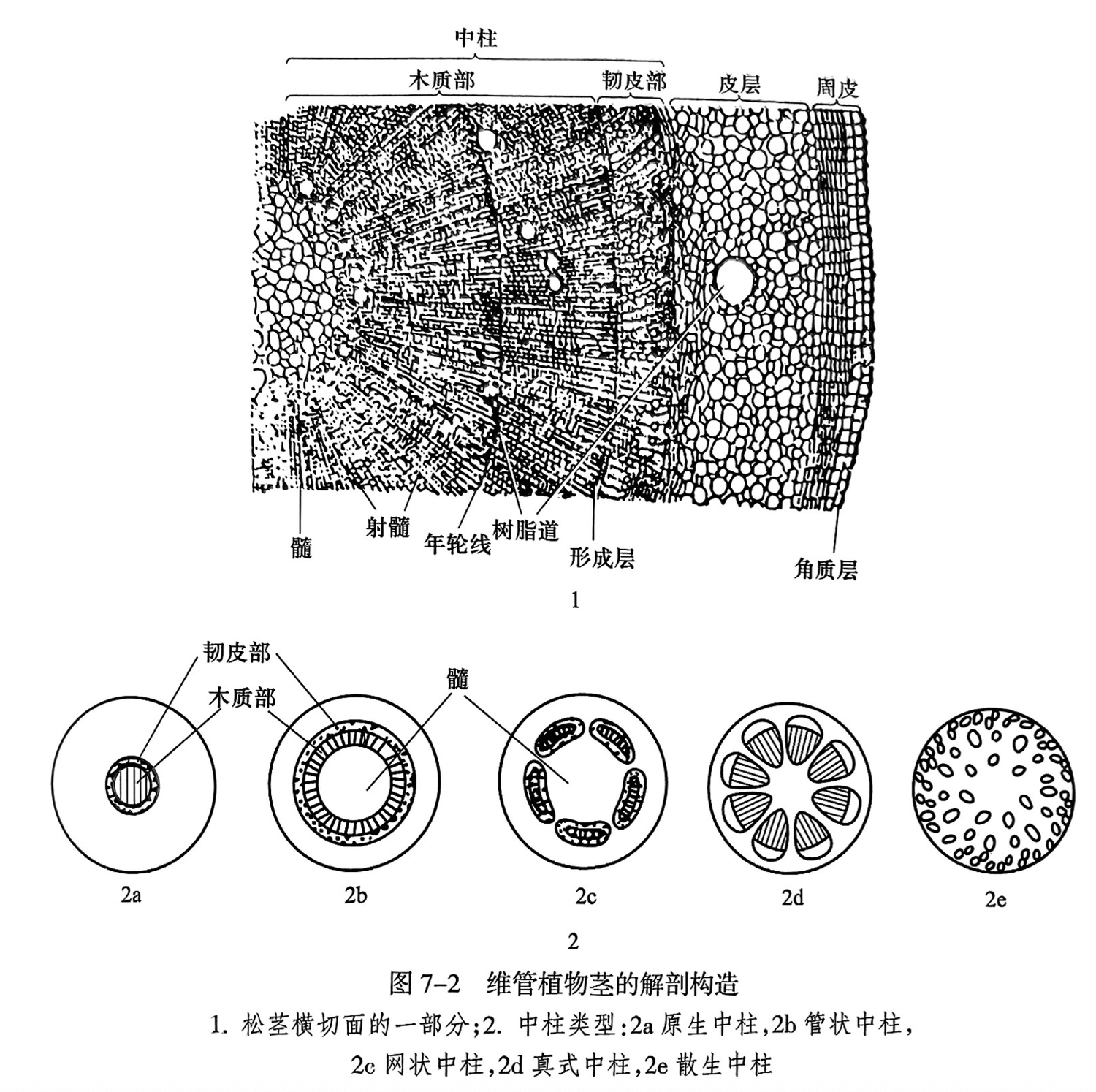
在地层中植物茎干化石十分常见,有些石化作用较好的木材化石可清晰地显示植物茎的解剖结构。现代各类高等植物茎的解剖结构差别较大,以裸子植物松的茎为例,可见茎的结构自外向内分为周皮(初生茎为表皮)、皮层和中柱(stele)三部分(图7-2之1)。其中,中柱是植物的重要的输导系统,主要由韧皮部、木质部和位于茎中心的髓(pith)组成。横向联结髓和皮层的薄壁细胞称为射髓(pith ray),呈辐射状。原始维管植物的茎中央无髓,称为原生中柱(protostele)(图7-2之2a)。原生中柱逐步演化为有髓、维管系统围绕髓的外面呈现圆管状的管状中柱(solenostele)(图7-2之2b)。管状中柱又演化出髓的外方为一圈大小不同彼此分离维管束的网状中柱(dictyostele)(图7-2之2c)。进化水平更高的是维管束被射线分隔的真式中柱(eustele)(图7-2之2d)和维管束分散在基本组织中的散生中柱(atactostele)(图7-2之2e)。因此,高等植物茎的中柱类型反映了它们的进化规律。
三、叶
叶(leaf)是制造有机物的营养器官,其主要功能是光合作用、蒸腾作用,还有一定的吸收作用和繁殖作用。叶由于数量多,表面有角质层保护,因而被掩埋后保存为化石的机会比较多。 在地史时期里,并不是所有植物都枝繁叶茂,早期原始植物无叶,之后才逐步进化出各种有叶梢物。无论是化石植物还是现代植物,叶的形状和叶脉的多样性在一定程度上反映了各植物种的特征,除少数异叶性情况外,同种植物的叶形相似。
(一)叶的组成
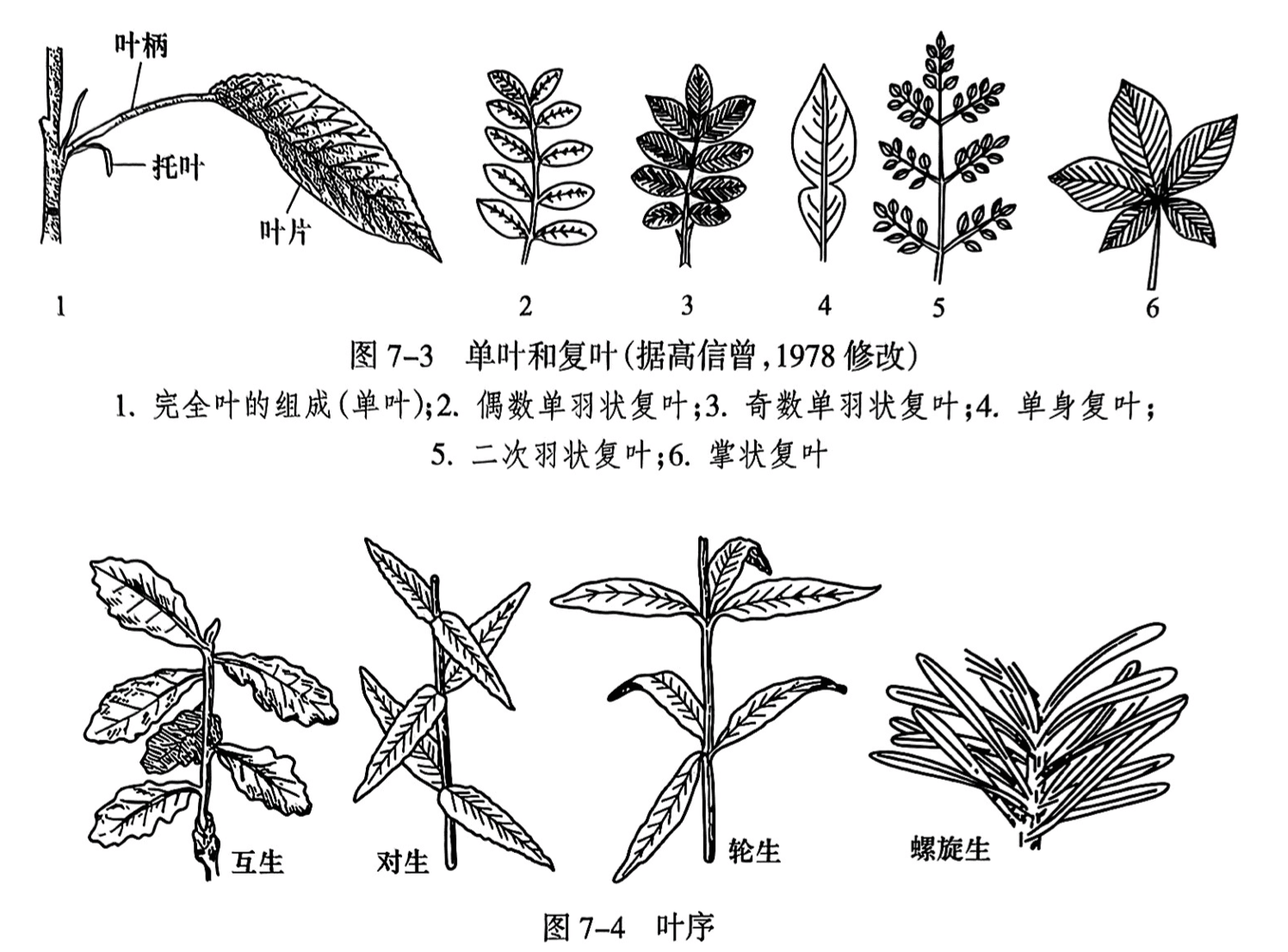
叶通常由叶柄(petiole)和叶片(leaf blade)组成,有的还有托叶(stipule)(图7-3之1)。没有叶柄的称为无柄叶(sessile leaf);叶柄上只有一枚叶片的称单叶(simple leaf);叶柄上有多片小叶的称为复叶(compound leaf)(图7-3);有的叶基部相连形成包围于茎节的叶鞘(leafsheath)。
(二)叶序
叶在枝上排列的方式称为叶序(phyllotaxy)。叶序有互生、对生、轮生、螺旋生等(图7-4)。叶排列的规律是使相邻叶之间互不遮盖,让叶片以较大面积接受阳光。
(三)叶的形状
叶的形状包括叶的整体轮廓、顶端、基部及边缘轮廓。叶的轮廓通常以叶的长、宽之比及最宽处的部位为标准而划分为基本的几何形态,并结合常见物体的形象而命名(图7-5)。

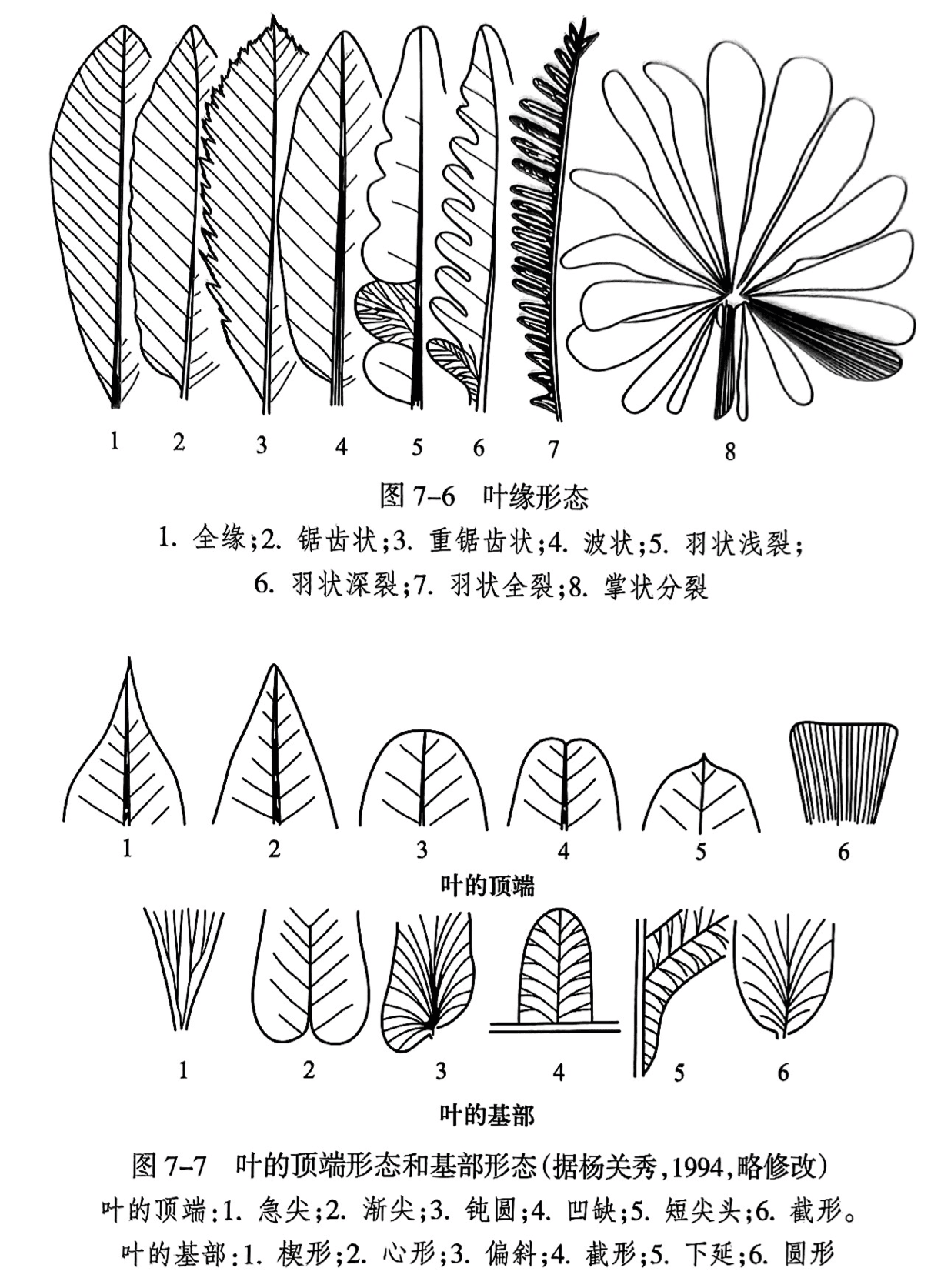
叶的边缘无裂者称为全缘,有的叶缘呈锯齿状、波状,或呈羽状浅裂、羽状深裂、羽状全裂和掌状分裂(图7-6)。叶的顶端形态和基部形态各有六种(图7-7)。
(四)脉序
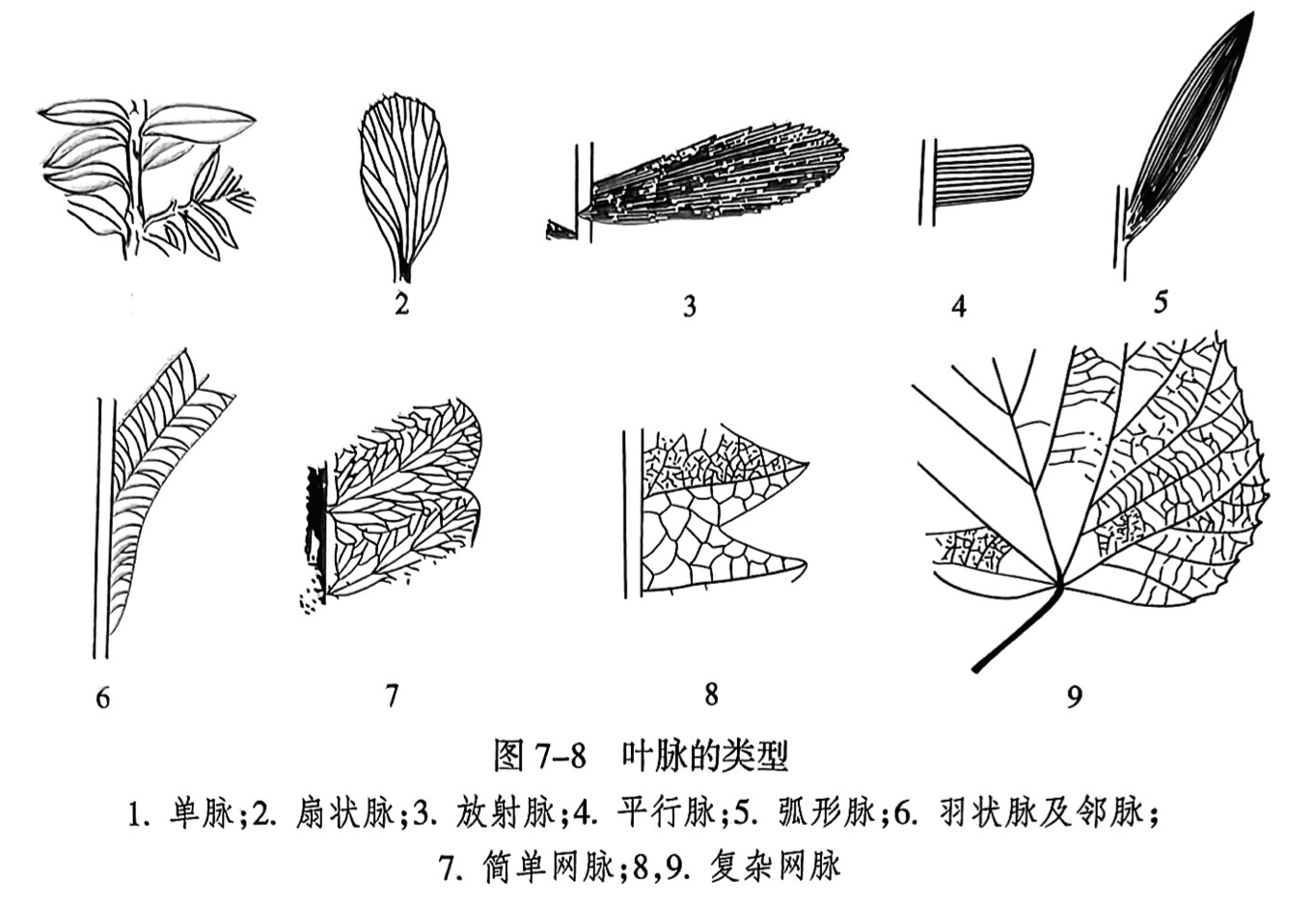
叶脉(vein)是分布于叶片中的维管束。叶脉在叶片中排列的方式称为脉序(venation),其形式多样,且比叶形更具有稳定性,是鉴定植物化石的重要特征,有8种基本类型(图7-8):
(1)单脉(simple vein)。叶片中只有一条叶脉自基部伸至顶端。
(2)扇状脉(flabellate vein)。叶脉均匀地几次二歧式分叉,呈扇状展布于叶面,如银杏。
(3)放射脉(radiate vein)。叶脉在叶基部多次二歧式分叉,在叶面较直地呈放射状伸出。
(4)平行脉(pararell vein)。叶脉在叶基部二歧式分叉,伸至叶面彼此平行。
(5)弧形脉(arch vein)。叶脉自基部伸出后,平行叶缘呈弧形至叶顶端汇合。
(6)羽状脉(pinnate vein)。有一条中脉或称主脉(midrib),两侧自中脉分出羽状排列的侧脉(lateral vein);侧脉也可分成几级,末级侧脉分叉或不分叉。有的侧脉不自中脉伸出,而出自羽轴称为邻脉。扇状脉至羽状脉都是开放脉序(open venation)。
(7) 网状脉(reticulate vein)。又称闭锁脉序(close venation),叶脉或侧脉二歧式分叉并相互联结成同一级别的单网者称为简单网脉或单网脉序(simple reticulate venation);复杂网脉(complex reticulate venation)的侧脉单轴式分枝结成网,网眼内又有细脉单轴式分枝组成次一级小网,依次可达3~4级网脉套叠,亦称重网脉序(double reticulate venation)。有的小网眼内还有不联结成网的细脉称盲脉(blind vein)。
(8)掌状脉(palmate vein)。叶内有几条等粗的脉(主脉),自基部呈辐射状伸出。
(五)叶的解剖结构
垂直叶片作切面可将叶分为表皮(epidermis)和叶肉(mesophyll)两部分,并且在切面上会有维管束(叶脉)贯穿(图7-9)。表皮覆盖整个叶片,通常分为上、下表皮。表皮是一层生活的细胞,不含叶绿体;这些细胞排列紧密,除气孔外无间隙,以防止水分蒸发。表皮细胞外壁角质化,并堆积成角质层,在干旱环境下角质层会增厚。表皮细胞间常有气孔器,由两个半月形的保卫细胞夹气孔组成;气孔可开合,在干旱环境下的气孔下陷。一般上表皮气孔器数量比下表皮少。有些植物化石经过处理,可显示清晰的表皮细胞及气孔器结构,是古植物系统分类和古环境分析的重要信息来源。叶肉由栅栏组织和海绵组织组成。栅栏组织(palisade parenchyma)位于上表皮下面,是排列比较紧密的长柱形细胞;海绵组织(spongy parenchyma)含大量的叶绿体,位于下表皮之上,是排列疏松的细胞(图7-9)。
(六)叶的生态类型
依照植物与水分的关系,可把植物分为旱生植物(xerophytes)、中生植物(mesophytes)和水生植物(hydrophytes)。旱生植物适应干旱环境;中生植物需要有充足的土壤水分和比较湿润的空气;水生植物则部分或全部植物体沉入水中生活,上述不同生态类型植物的叶的形态和结构具有较大差异,具体如下。
(1)旱生植物的叶。旱生植物叶小而厚,或多细毛,叶的表皮细胞壁厚,角质层发达。有些旱生植物,表皮由多层细胞组成,气孔下陷。旱生植物的另一种类型,称为肉质植物,例如,龙舌兰、芦荟、景天、马齿苋等,它们的一个共同特征是叶肥厚多汁,叶内有发达的薄壁组织,蓄存大量水分。
(2)中生植物的叶。它们的形态和结构介于旱生植物的叶和水生植物的叶之间。
(3)水生植物的叶。水生植物的叶片薄,有的叶片分裂成丝状,有利于增大叶片与水的接触面积,叶的表皮未发生角质化,没有角质层或角质层很薄,细胞内含有叶绿体,叶表皮没有气孔,叶肉没有栅栏组织和海绵组织的分化,但有发达的细胞间隙组成的通气道。维管组织退化(特别是木质部),机械组织不发达。
四、植物的繁殖与生殖器官
1.植物的繁殖
植物的繁殖方式可分为营养繁殖、无性生殖和有性生殖三种类型。营养繁殖(vegetative propagation)是植物营养体的一部分从母体分离开(在有些情况下不分离开)直接形成新个体的繁殖方式,如葡萄的扦枝,土豆块茎芽的分殖等。无性生殖(asexual reproduction)是植物的营养体生长到一定时期,进入生殖生长阶段,产生具有生殖功能的细胞,这些细胞不经两性的结合可直接发育成新个体。这种具有生殖功能的细胞称为孢子(spore),因而无性生殖也称为孢子生殖(spore reproduction)。有性生殖(sexual reproduction)是植物生长到某个阶段,形成有性细胞,这种有性细胞称为配子(gamete);由雌、雄配子结合成合子(zygote),合子萌芽形成新植物体。最广泛而高级的有性生殖是卵式生殖(oogamy),其雌、雄配子的形状、大小、特征差异很大,雄配子(精子)极小,有鞭毛,活动积极,但在种子植物中,精子无鞭毛,随原生质流动;雌配子(卵细胞)大,无鞭毛,不活动,位于颈卵器内(保护卵细胞的瓶状器官)。
2.生殖器官及相关化石
生殖器官是古植物分类和鉴定的重要依据,它的形态和结构在不同植物类别中差别很大。此外,生殖器官在不同发育阶段可以形成不同类型的化石,特别是生殖器官在发育过程中产生的孢子、花粉常形成量多、分布广的微体化石。
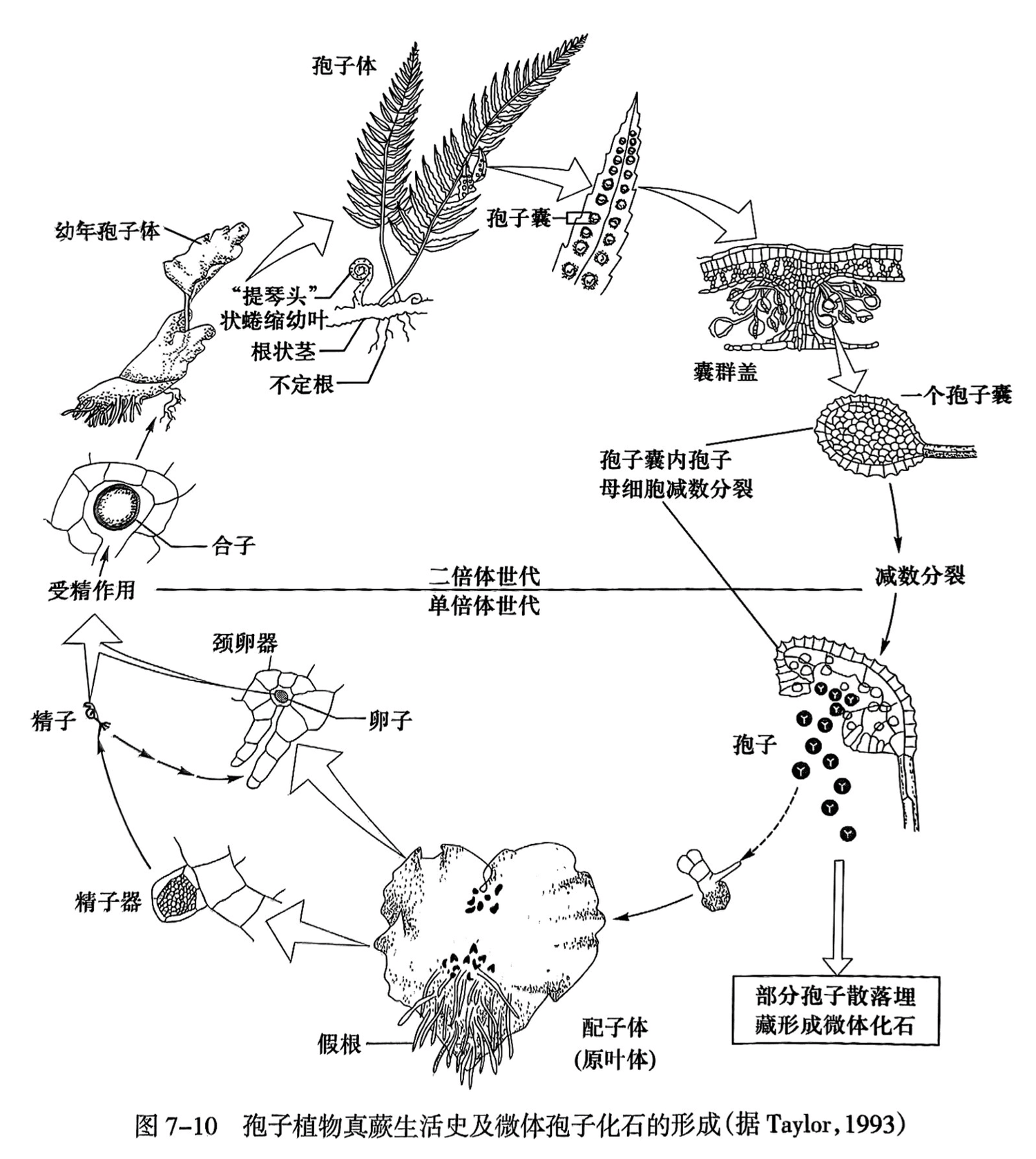
苔藓及蕨类植物的无性生殖器官是孢子囊(sporangium),孢子囊发育成熟形成孢子,由孢子发育成新的植物体。孢子埋藏常形成大量的微体化石(图7-10)。苔藓及蕨类的有性生殖器官形成的化石比较少见,它由多个细胞构成:雄性生殖器官称为精子器,其内产生精子;雌性生殖器官称为颈卵器,其内产生卵细胞。精子成熟后,精子器破裂,精子溢出,以水为媒介,游进颈卵器内,使卵细胞受精成为合子;由合子发育成胚,再由胚发育成新的孢子体(图7-10)。
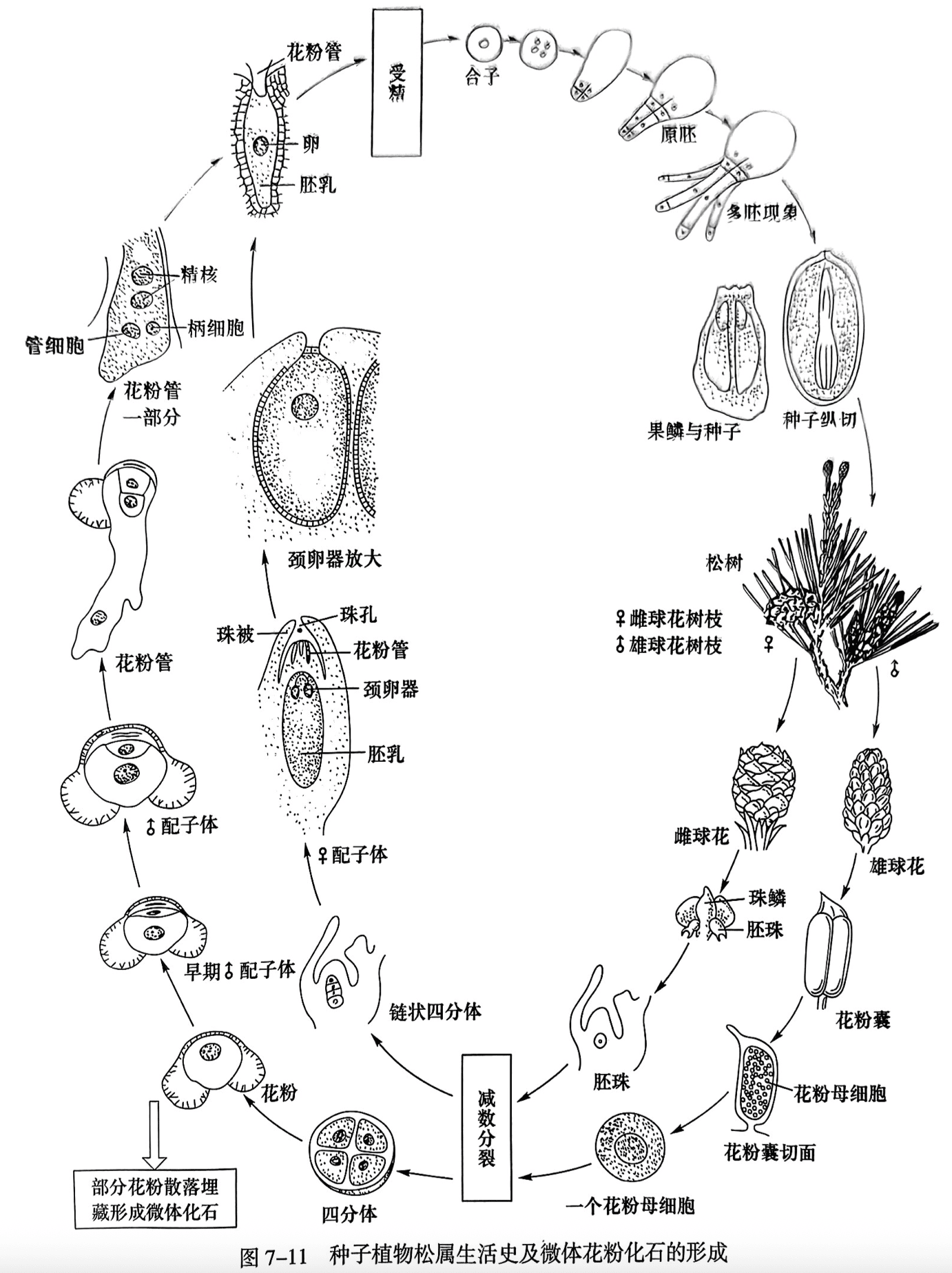
裸子植物的生殖器官是球花(cone)或称孢子叶球(strobilus),它的结构与蕨类植物的生殖器官有同源关系。球花通常都是单性,有雄球花(male cone)和雌球花(female cone)之分,雄球花和雌球花或同株或异株。雄球花(小孢子叶球)即由雄蕊(小孢子叶)聚生而成,雄蕊下面生有花粉囊(小孢子囊)。内有多个花粉母细胞(小孢子母细胞),花粉母细胞经减数分裂产生花粉,再由花粉粒发育成雄配子体。花粉粒经过埋藏也可形成大量微体化石(图 7-11)。雌球花(大孢子叶球)生有一至多个裸露的胚珠.胚珠经减数分裂产生雌配子体,雌配子体的近珠端产生颈卵器,花粉粒终风传播在胚珠的珠心上萌发形成花粉管进入胚囊,产生精子与颈卵器内的卵细胞结合成为合子,经过多胚阶段发育成种子,再由种子萌发成新的植物体。
被子植物的有性生殖器官是花朵(flower)。花朵由花柄、花托、花萼、花冠、雄蕊群、雌蕊群几个部分所组成、雄蕊的花粉囊产生花粉,这些花粉埋藏亦可形成大量的微体化石。雌蕊的子房内包藏着胚珠。胚珠由珠柄、珠被、珠孔和胚囊构成;胚囊包含8个核,其中的一个核发育成卵细胞、花粉形成后。通过传媒到达雌蕊:花粉进一步发育产生的雄配子(精子)进入胚囊,与卵细胞结合形成合子。由合子发育成胚,同时由胚珠发育成种子,再由种子萌发成新的植物体。
3.世代交替
植物在生活史中出现了两种个体。一种是能产生配子,行有性生殖的配子体(gametophyte)。配子体是由孢子发育形成的,为单倍体。另一种是能产生孢子,行无性生殖的孢子体(sporophyte)。孢子体是由合子发育形成的,为二倍体。我们把第一个阶段称为配子体世代,或称有性世代;把第二个阶段称为孢子体世代,亦称无性世代。在植物的生长发育过程中,两个世代有规律地交替出现,故称为世代交替(alternation ofgenerations)。但各类植物发育情况不同,例如,蕨类植物的无性世代和有性世代彼此独立,以无性世代的孢子体占优势,产生大量孢子繁衍后代;种子植物则世代交替不明显,配子体退化,寄生在孢子体上,有性生殖结果产生种子繁殖后代。