3 软体动物
内容节选自《古生物学》(童金南主编,2021年版)第五章 古无脊椎动物 第三节 软体动物
一、 一般特征
软体动物(mollusk)是无脊椎动物中数目、种类众多的一个门类,已知的现生种和化石种~共有12万余种. 是动物界中仅次于节肢动物的第二大门类。它们分布广泛,适应能力强. 在陆上、淡水、咸水中均有代表. 如蜗牛、河蚌、海螺、乌贼等。各类软体动物虽然形态习性差异较大,但基本特征相似. 如身体柔软而不分节,一般可分为头、足、内脏团和外套膜4部分。具口的头部位于身体的前端. 但其在不同的类别中发育程度有所差异。除双壳类外,其他各类软体动物的口腔内有颚片和齿舌。足部具有发达的肌肉,常位于腹部,用于行动,但随生活方式的不同而有各种不同的形状。内脏团是各种内部器官所在之处,为动物的躯体部分。外套膜包裹着内脏团. 常分泌钙质的硬壳,用以保护身体。外套膜形状随种类而异,所形成的硬壳形状也各不相同。除大多数成年期的腹足纲软体动物外,软体动物的壳体均为左右或两侧对称(图5-10)。软体动物的呼吸器官为鳃或肺,水生者用鳃,陆生者用肺。其繁殖方式一般为卵生,也有胎生。
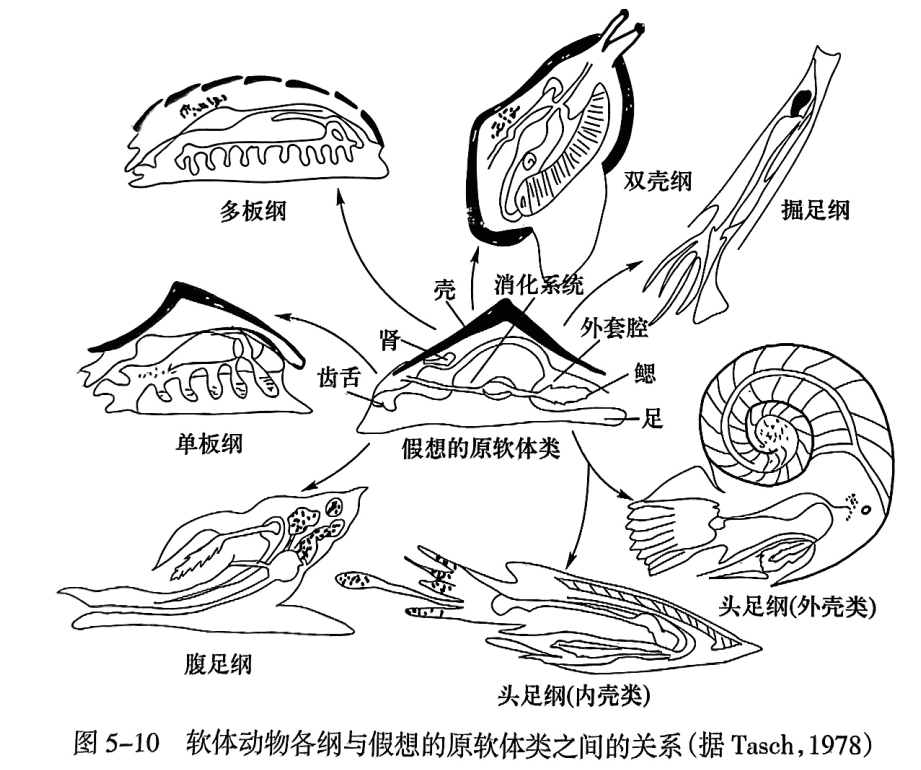
根据硬壳和软体等特征,软体动物门可分为10个纲,即单板纲(Monoplacophora)、多板纲(Amphineura)、无板纲(Aplacophora)、喙壳纲(Rostroconchia)、掘足纲(Scaphopoda)、腹足纲(Gastropoda)、双壳纲(Bivalvia)、头足纲(Cephalopoda)、竹节石纲(Tentaculita)和软舌螺纲(Hyolitha)。
二、双壳纲
(一)一般特征
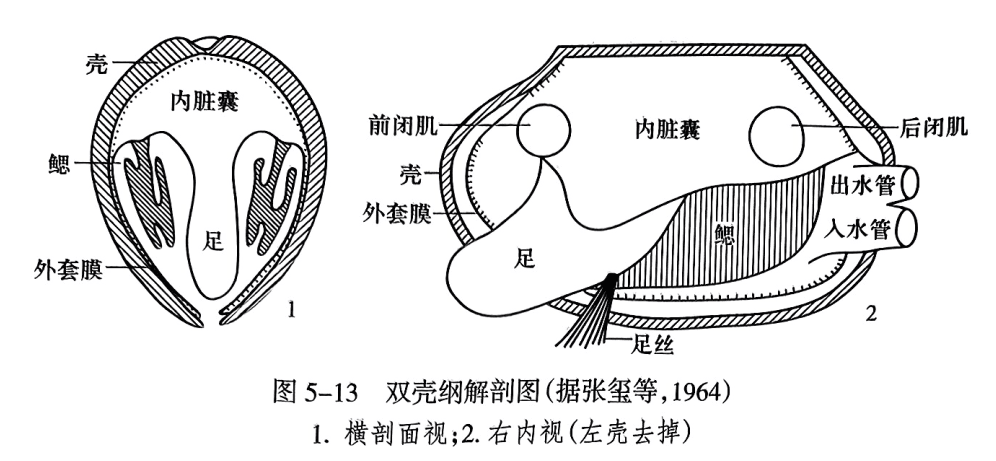
双壳纲(Bivalvia)是水生软体动物,绝大多数为两侧对称,常见的有壳菜蛤、蚶、河蚌等。其身体具有由包裹内脏团的左右两片外套膜(mantle)分泌出的一对起保护作用的钙质外壳,故被称为双壳纲(Bivalvia Linnaus,1758)。外套膜之间的外套腔(mantle cavity)内生有起呼吸作用的瓣状鳃,故也被命名为瓣鳃纲(Lamellibranchiata de Blainville,1824)。足位于身体前腹方,常似斧形,故又名斧足纲(Pelecypoda Goldfuss,1820)。足伸出于两壳瓣之间,用以挖掘泥沙、移动身体或钻孔等。很多双壳纲还在足后伸出一簇丝状的足丝(byssus),以黏着在外物上(图5-13)。有些成年个体足丝发育,而足退化。双壳纲头部退化,又有无头纲(Acephala Cuvier,1798)的名称。根据优先律,Bivalvia命名最早,因此目前一般都使用“双壳纲”一词。
双壳动物依靠外套膜上的纤毛有规律地运动,形成出、入两股水流。入水流带来食物和氧气,出水流排出新陈代谢废物。有些种类的后部外套膜边缘愈合并向后延伸,分别形成入水管(inhalant siphon)(在下)和出水管(exhalant siphon)(在上),以分开出、入水流(图5-13之2);一些深埋穴居的双壳纲,其水管还会向后伸长,以到达沉积物表面,进行水流交换。
(二)基本构造
1. 壳形
双壳纲一般具有两个互相对称的、大小一致的两瓣壳(左壳和右壳)。每瓣壳本身前后一般不对称,其上、下、前、后边缘分别称作背缘、腹缘、前缘和后缘。有些种类由于对固着、漂游或偃卧生活的长期适应,造成两瓣不等。双壳纲常见的壳形见图5-14。

2. 壳的外部结构
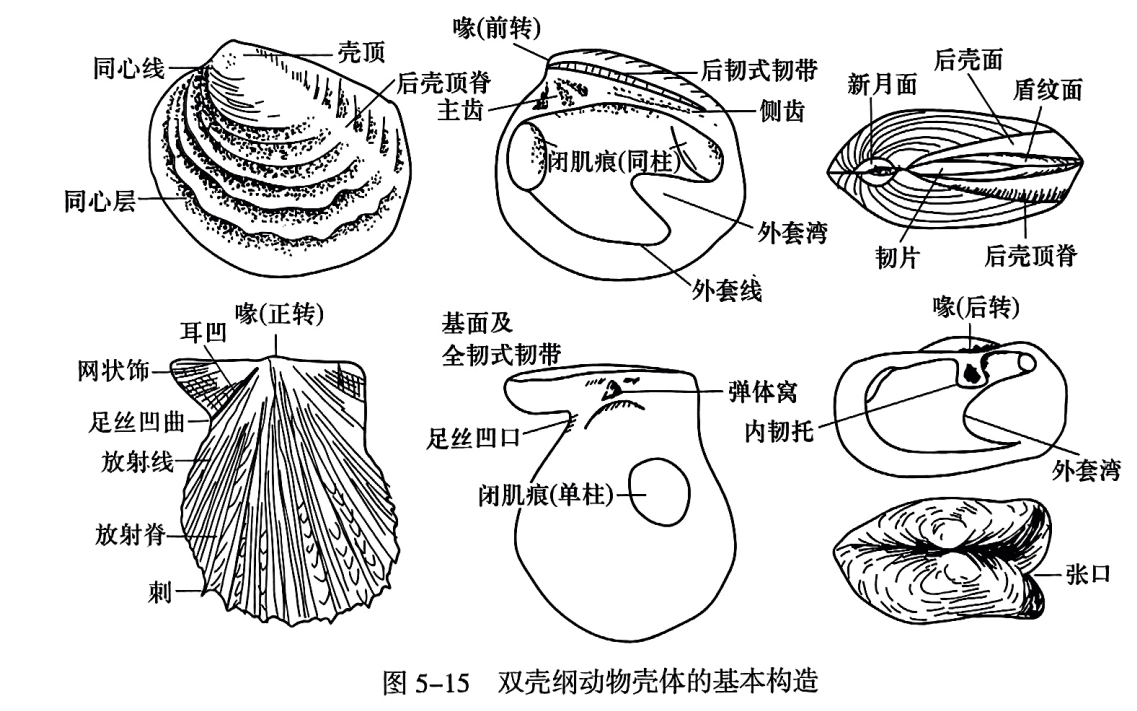
双壳纲壳体最早形成的壳尖称为喙(beak,或称壳嘴)。喙多数向前指(称为前转,prosogyrate),也有在垂直面上的(正转, orthogyrate)或向后指(后转, opisthogyrate)者。包括喙在内的壳顶部最大弯曲区称壳顶(umbo)。有些种类由喙向后腹方伸出一条隆脊,称后壳顶脊(posterior umbonal ridge)。在喙下常有一个在成因和形态上均与壳面不同的面,是连接两瓣壳并协助两壳打开的弹性有机质韧带(ligamant)的附生处。有的位于喙与两壳铰合线之间,或平或凹,称为基面(cardinal area);有的位于喙前呈心脏形凹陷,称新月面(lunule);有的位于喙后呈长槽形凹陷,称盾纹面(escutcheon);新月面与盾纹面可以并存。有些种类的壳面在喙的前侧或后侧变得扁平,并呈翼状伸出,称为前耳(anterior auricle)、后耳(posterior auricle)或后翼(posteriorwing)。它们与其余壳面或呈过渡状态,或以槽状凹陷隔开,此凹陷称为耳凹(auricular sulcus)。有些类别在前耳的下方有足丝伸出,因而在前耳凹与前缘相交处形成一个缺口,并在缺口的壳内侧留下足丝穿过的齿状痕迹————丝梳(ctenolium)。该缺口通常在右瓣上较深,称为足丝凹口(byssal notch);在左瓣上较浅,称为足丝凹曲(byssal sulcus)。有些营深埋穴居生活的类别,当其水管不再能够收回壳体内部时,则在壳体的后方保留一个张口(gape)(图5-15)。
3. 壳饰
双壳纲壳体表面除少数光滑者外,通常具有发育程度不等的装饰。它们的壳面装饰大体可分为同心和放射两类,每类又各按强度分为纹、线、脊、褶(或层)等。同心装饰反映了生长过程,也称为生长纹(最细一级)、线、脊、层等。放射纹从壳顶向周缘散开,一方面会逐步增粗和变得稀疏,另一方面会逐步增多。放射脊增多的方式主要有两种,即分叉式和插入式。前者是指放射脊一分为二或一条分为多条(呈束状);后者是在脊间的放射沟中生出新的放射脊。有的种类同时具有以上两类壳饰,二者相交成网状(图5-15)。有的还具有瘤、节或刺。壳体内面通常是光滑的,但有时也有与壳表面相对应的纹饰,甚至具有与表面不同的纹饰。
4. 壳的内部构造
壳的内部构造基本上可分为4类:外套膜附着痕、肌肉附着痕、韧带附着痕和铰合构造(齿系)。
(1)外套膜附着痕。指外套膜外缘游离部分与其在壳内面附着部分之间在壳内留下的痕迹,也称为外套线(pallial line)。它与腹缘大致平行,在背部左右两瓣外套膜互相连接。具有水管的壳,当双瓣关闭以御敌或阻止泥沙进入时,须将水管拉入壳内,外套膜附着痕因此向内移动,使外套线形成弯曲,称外套湾(pallial sinus)。在海底表面或浅埋生活的双壳纲中通常没有长的水管,其外套线上没有外套湾。钻入海底泥沙或岩石中生活的种类,如果有可伸缩的水管,水管越长则收入壳内时水管占用的空间越多,外套湾也就越深。少数种类水管不再收入或不完全收入壳内,在壳上留下张口,其外套湾则不一定很深(图5-15)。
(2)肌肉附着痕。这主要指由使壳体闭合的闭壳肌(adductor)所产生的闭肌痕(adductorscar)。双壳纲一般有1~2个闭壳肌,但由于适应不同的生活方式,其大小发生改变,因而形成了不同的闭肌痕类型。此外,双壳纲还有伸、缩肉足的肌肉所留下的足肌痕(pedal scar),它们常位于前闭肌痕的上方或下方。
(3)韧带附着痕。指司壳体开启的韧带附着所留下的痕迹。韧带(ligament)按物质成分分为外层的薄片质(受拉张时产生收缩压力,也能承受压力)和内层的纤维质(受压缩时产生弹性张应力. 不能承受张力)。按其在壳上的位置可将其分为位于壳外的外韧带(external ligament)和位于壳内的内韧带(international ligament)。韧带附着物主要有4种:①基面(cardinal area)、新月面和盾纹面,为外韧带附着处;②韧片(nymph),是喙后沿铰缘分布的狭长片状板,绝大多数为外韧带:③弹体窝(resilifer),为喙下的三角槽,一至数个,是纤维质弹回体(resilium, resilia)(壳内或壳外)的附着处;④内韧托(chondrophore),是由喙下向前伸出的凹板,附着弹回体。
双壳纲壳体的开合就是借韧带与闭肌配合进行的,即以铰合构造为支点,当闭壳肌收缩时,具有弹性的外韧带被拉伸,壳体便关闭;当闭壳肌松弛后,外韧带韧性收缩而使壳体开启。内韧带弹回体主要是以压弹性协助外韧带将壳体弹开。
(4)齿系(dentition)。指的是两瓣壳之间的壳体铰合构造,其在壳体构造中具有最显著的分类意义和化石鉴定意义。它位于铰缘之下,作为两瓣的关节支点,由铰齿(tooth)及铰窝(socket)组成,通常位于沿铰缘分布的铰板(hinge plate)上。与腕足纲不同的是:双壳纲每一瓣上铰齿与铰窝相间排列,且与另一瓣上间列的铰窝与铰齿相对应,两者相互嵌合。齿在演化中分异为主齿和侧齿。主齿(cardinal tooth)位于喙下,较粗短,与铰缘呈较大角度相交。侧齿(lateral tooth)远离喙,片状,与铰缘近于平行。铰齿和铰窝的种类、数目、形状和排列的总体情况称为齿型(dentition),主要有以下几种类型(图5-16):
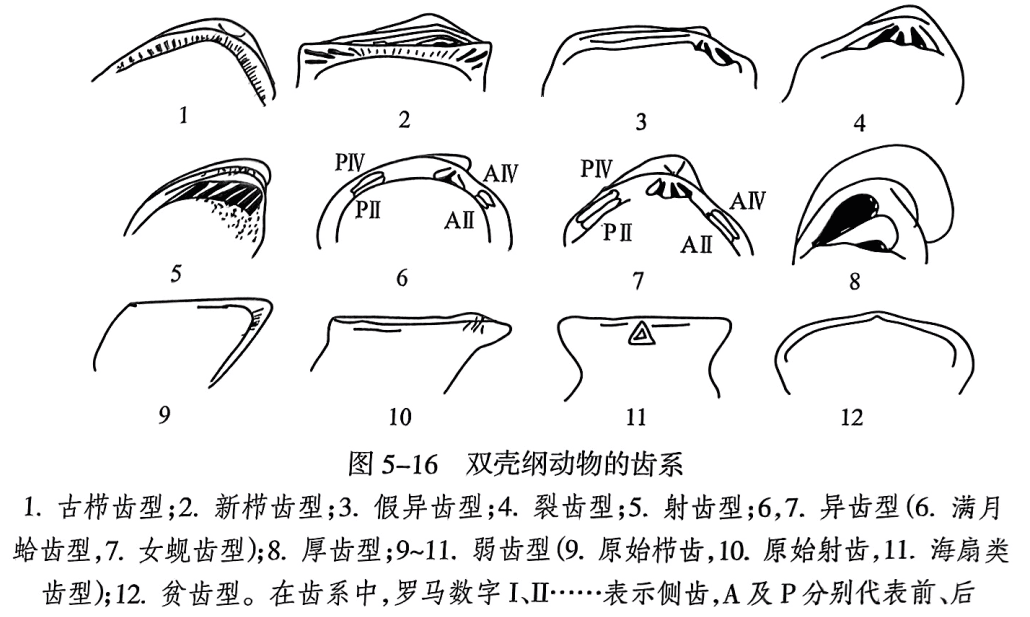
栉齿型(taxodont):铰齿多,短而小、排列与铰缘垂直或倾斜. 倾斜方向往往向腹方集中。原始类型无基面,称古栉齿型(palacestaxodent);进步类型有基面,称新栉齿型(tietaxrelent)。
异齿型(heterodont);铰齿分化为中央(壳喙下)的两个或三个粗短的主齿(cardinal teeth)及前、后侧远离壳喙的狭长片状侧齿(lateral teeth),前、后侧齿至多各两个,与铰缘平行。
古异齿型(palaecheteredent):是介于栉齿型与异齿型之间的一些过渡类型。如由喙向下放射分布的一系列铰齿构成的射齿型(actiondont),代表了在演化过程中铰齿数减少,强度增大,最后每瓣仅在壳喙之下留下两至三个粗长的主齿的现象。如果其中居中的主齿下部发生分裂,在左瓣上为“人”字形,右瓣上为“八”字形,就将这种齿系称作裂齿型(schizodont)。另一种情形是,每瓣有一至三个铰齿沿铰缘向前、后延长呈片状。这种类型是向前的齿较短. 看似主齿,向后的齿长,形态上很像异齿型中的侧齿,但其起自喙下,故称为假侧齿或片状齿(lamelliform teeth)。相应地,前方的主齿也只能称为假主齿,这种齿系称为假异齿型(pseudoheterodont)。
厚齿型(pachyodont):由异齿型向固着底栖方向演变而成的一种特殊齿型。这种类型的铰齿粗厚巨大,一瓣两个,另一瓣一个。
弱齿型(dysodont):这一类的铰齿始终处于萌芽或不发育阶段,包括原始栉齿式、原始射齿式,或沿铰缘对称地微呈齿片状(见于海扇类,称为铰棱)。弱齿型与幼年或终生以足丝固着有因果关系,为翼形亚纲所特有。
等齿型(isodont):这种类型每一壳瓣具有两个发育程度大致相等的铰齿,分别位于弹体窝的前、后,与生物以壳体固着生活有关。
贫齿型(desmodont):由于钻孔穴居,这种类型的生物的铰齿全部消失,或仅有一枚弱齿,一般具有内韧托和外套湾。此外,古生代还有一些非穴居的无齿种类,称作隐齿类(cryptodonts)。
5. 壳的定向与度量
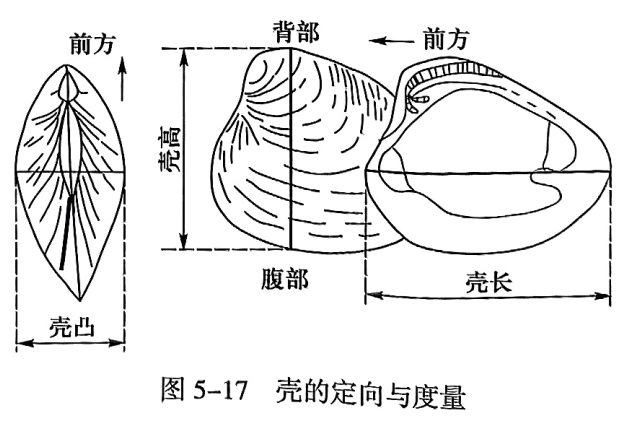
双壳纲为两侧对称动物,因此其壳体分为前、后、背、腹、左、右。进行壳体定向时先根据下列各条规则确定前后(图5-17):
(1)喙一般指向前方(少数相反,如三角蛤类)。
(2)放射及同心装饰一般由喙向后腹方扩散。
(3)壳体前后不对称者,一般后部较前部长(少数相反,如栗蛤类)。
(4)新月面在前,盾纹面在后。
(5)足丝凹缺(凹口)在前,后耳常大于前耳。
(6)外套湾位于后部。
(7)两条闭肌痕不等大时,大者在后;只有一条闭肌痕时,一般位于中央偏后部。
先定前后,再定背腹及左右,即将两瓣的铰合部设置为向上,开闭部设置为向下,并将壳的前方指向观察者的前方。如此拿定,则上方为背,下方为腹;左侧的壳瓣为左壳,右侧为右壳。
壳的度量:前后两端最大长度距离为壳长;由壳顶至腹缘垂直壳长的最大高度为壳高;垂直于两壳接合面的最大距离为凸度;单壳内表面与外表面之间的厚度为壳层厚度,各点壳厚度不一。
(三)双壳纲的分类
双壳纲的分类是以齿系为主要根据,并综合其他特征(鳃、外套膜、足丝、闭肌痕及壳质成分等),共分为6个亚纲(表5-5)。
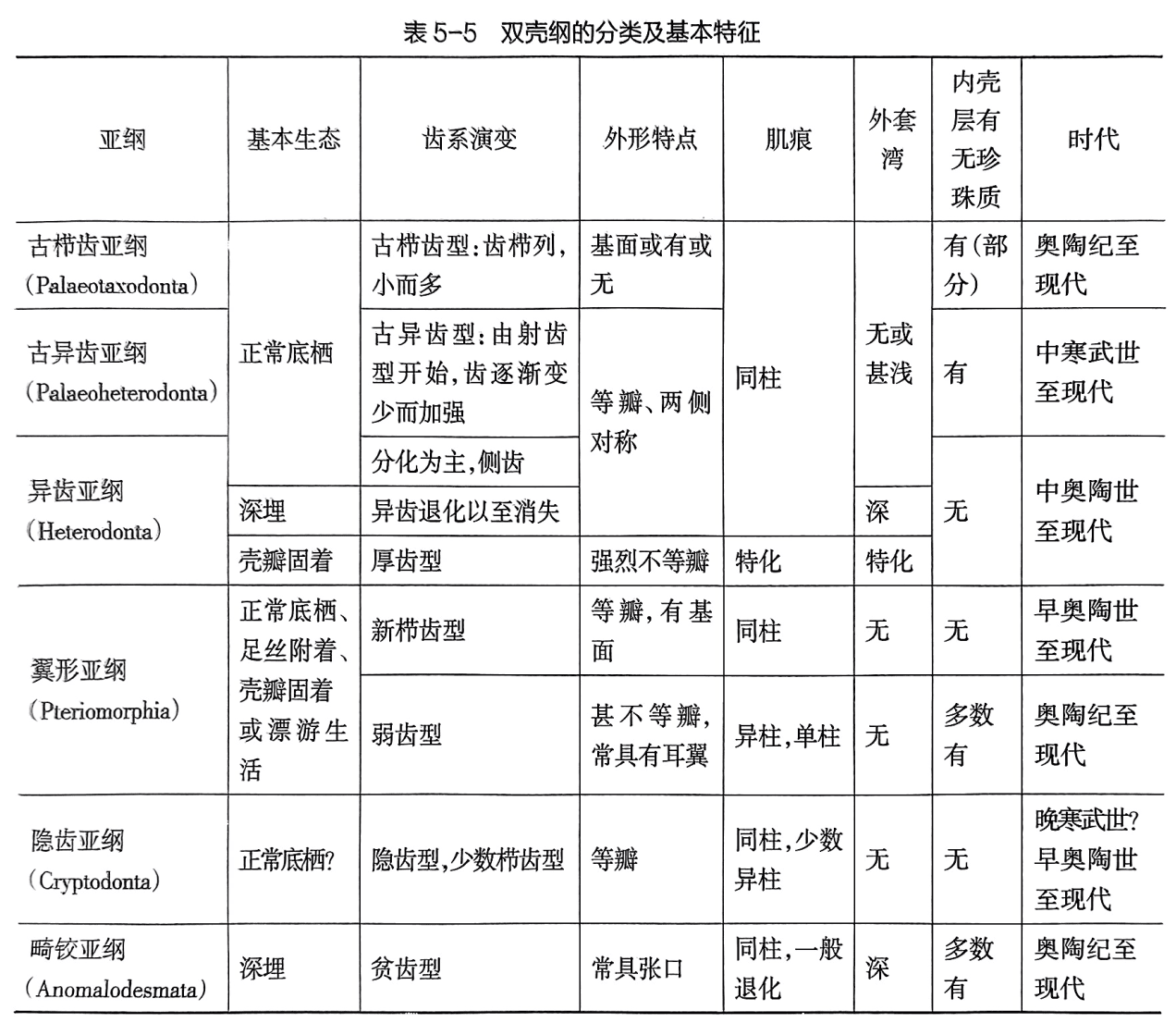
(四)双壳动物的生态及其与硬体结构的关系
双壳动物的生活方式十分复杂多样,如图5-18所示。从演化来说,有三类基本的或原始的生活方式——正常底栖、足丝附着、深埋穴居。三类生活方式长期地适应演化,在壳体形态、齿系、闭壳肌、足丝、水管等各方面都发生了相应的适应性变化,便形成各具特色的不同类别(亚纲和目)。
第一类为正常底栖(这是向足丝附着过渡的一类),通常水管不发育,少数在海底爬行(图5-18之10,13),多数浅埋入泥沙中(图5-18之1,11,12,14)。这一类壳体两瓣等大,且前、后差异不大,因此闭肌常为同柱;经常使用足,没有足丝及相关构造———足丝凹缺、耳等(部分箱蚶目除外);由于埋入得不深,适应抗御风浪、急流、敌害等,因而铰合及开闭机构(闭肌、韧带、齿系)发达;铰齿的演化方向是:强度加强,主、侧齿分化,同时齿数减少,结果由栉齿型及射齿型演化为古异齿型,再演化为异齿型;外套湾不存在(爬行和半身埋入的)或较浅(全身埋入,具明显水管)。这一类有辐射适应,例如,某些类别改营深埋穴居(图5-18之15,24,25,26)和壳瓣固着生活(图5-18之17),从而分别产生了与下述第二类、第三类相似的趋同变异(如图5-18之24,25,26的体形延长,外套湾深,铰齿退化,与第三类趋同)。其中固着生活的类别进一步演化,适应于在热带礁区以壳体的一瓣固着生活(图5-18之18,19),并随之产生一系列高度特化的性状——厚齿、厚壳、极不等壳、特化的肌痕和水管区。
第二类是以足丝附着生活,其暂时或永久地附着于水中物体上(图5-18之3,4,8),足丝向前腹方伸出,使足和壳的前部退化;壳形显著不等侧(飘形、翼蛤形等);闭肌亦随之由同柱演化为异柱,再演化为单柱;相关地,与足丝有关的构造(耳、足丝凹缺等)比较发育;由于在水底表面生活,故无外套湾。这一类后来向着偃卧(图5-18之6)、隙居筑巢(图5-18之5)、漂游(图5-18之7)或壳瓣固着(图5-18之9)等方向辐射适应;偃卧和漂浮者形成次生的侧扁壳形(图5-18之6. 7),但仍遗传保留了足丝附着时期的特点(耳、单柱等);壳瓣固着者(图5-18之9)产生与图5-18之17,18,19相似的趋同变异,如强烈不等壳。这一类在表生生活中经常开闭双壳,没有形成坚固铰合的铰齿,其开闭铰合的作用部分地被发达的韧带所代替,故这一类属于翼蛤目与壳菜目。
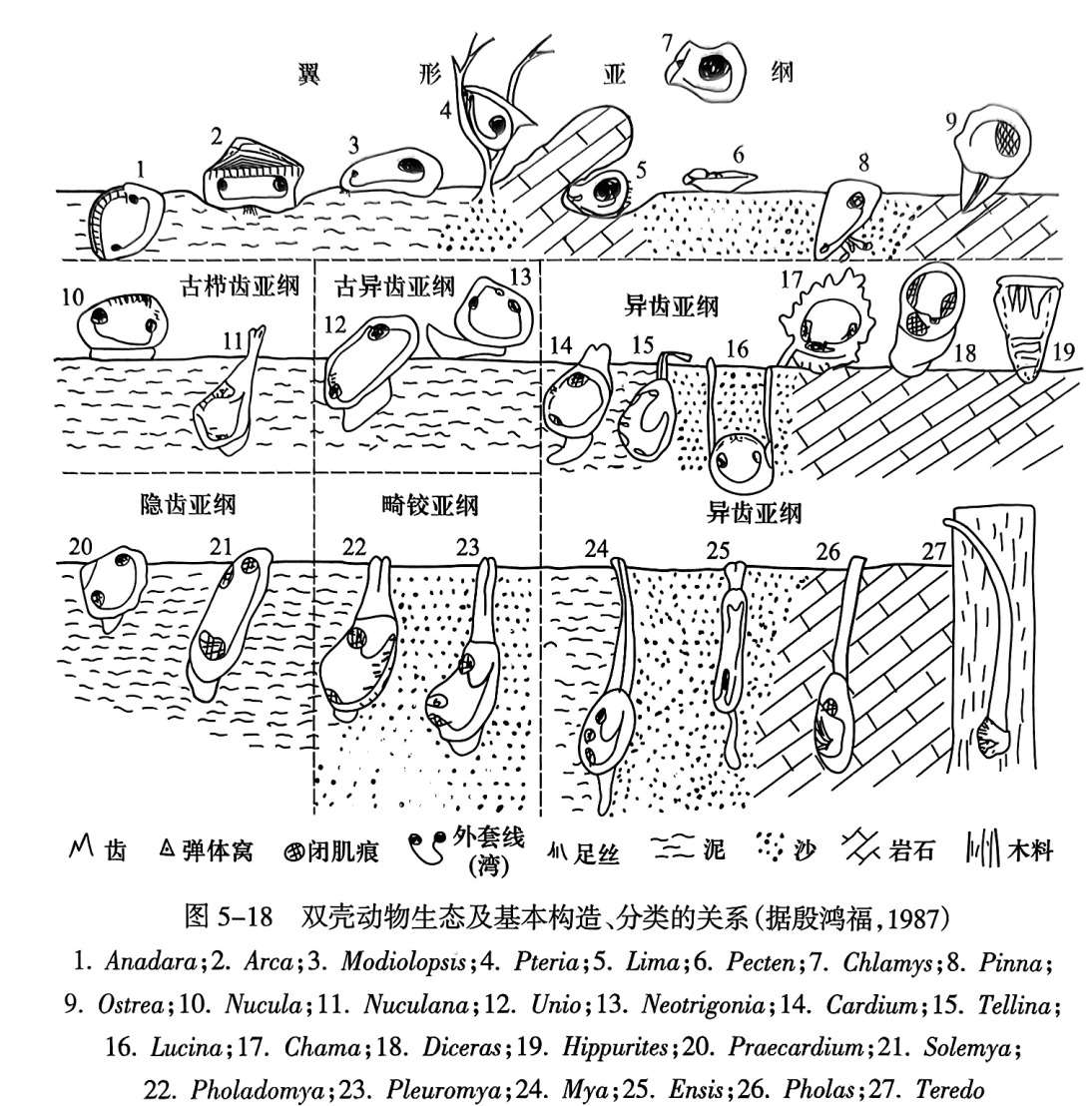
第三类为深埋穴居,是在泥沙中挖掘深的洞穴,长期或永久地穴居(图5-18之22,23)。这类生物的身体伸长. 喙不突出,外套湾深:有些由于水管和(或)足不再缩回而形成前、后张口;开闭壳体的齿系和闭肌退化. 铰齿弯消失。而适应于岩石钻孔(图5-18之26)、凿木(图5-18之27)和偃卧等生活的类别也各有一些新的特点,如钻孔者有锉刀状的前部和能使锉状前部往复锉动的肌肉-韧带系统等。
综上可见、双壳纲动物的生态与壳体构造具有密切的内在联系,而壳体特征又是双壳纲分类和演化的直接证据。这种联系简要概括在表5-5中,每一目的各种构造特点(壳形、齿系、肌痕、外套湾等)都是相关地配合而不是孤立形成的,每一套相关的构造又都是在一定的生态适应中长期演化的结果。其中有些是本身的适应变异,有的是祖先类型适应变异的遗传(亦往往重现于个体发育过程中)。但是. 实际情况要比上述内容更错综复杂,不能简单对待,以一概全。
(五)双壳纲的地史分布
已知双壳纲的最早代表(Fordilla)出现于早寒武世。双壳纲总的演化趋势是自显生宙以来逐步发展. 特别是从中生代开始迅速发展,现在达于全盛。其中具备水管的类型比不具备水管的类型增长远为迅速,其科数达后者之3倍,说明本纲的成功主要是由于穴居。在双壳纲的整个演化过程中有4次重大转折点:①奥陶纪初的辐射演化,由寒武纪17属34种猛增至奥陶纪199属1419种(1971资料),所有主要类别均已有代表。②古、中生代之交的大灭绝(60%的属和95%以上的种灭绝),但继之发生了高速的辐射演化。③三叠纪末的大灭绝(42%的属和90%以上的种灭绝),但继以新的辐射演化。④中、新生代之交的大灭绝(马尾蛤目、大部分三角蛤类和叠瓦蛤类灭绝),再次继以新的辐射演化。双壳纲的演化图谱是一个由几个间断期和相对平衡期交替组成的总体呈上升趋势的曲折线。
在本纲的演化过程中还有两个重要现象值得注意,第一个现象是双壳纲与腕足纲在地史过程中互为消长。这两类生物都是正常浅海底栖滤食性动物,因此其关系是相互竞争的。从内因来看,双壳纲比腕足纲优越(前者有较高级的器官,对盐度变化的适应能力强,有自由游泳独立摄食的幼虫),因此竞争结果是双壳纲成为最终的胜利者。但是,由于腕足纲出现得早(前寒武纪末期),当双壳纲低级代表出现时,腕足纲已经发展成熟了,其生理机能充分适应了当时的海洋环境,因而占据了古生代浅海等有利生活的领域;而这时的双壳纲被排挤在较深的水域和内海等环境中。只是经过相当长的历史时期,双壳纲逐渐向高级发展(器官的完善、壳的加固、铰合构造的发展等),它才逐渐击败竞争对手。外因(加里东和海西运动引起的重大环境变化)也起了促进作用。在泥盆纪时,双壳纲取得的第一个胜利是逐渐在海洋中扩展,并进入淡水环境。但这两纲生物的重大转化发生于古、中生代之交,二叠纪、三叠纪之交的大灭绝事件之后,腕足纲基本上退出了浅海生态环境,从三叠纪起双壳纲取得了绝对的优势地位。
第二个现象是双壳纲占领淡水领域,它在不利环境中逐渐产生了对广盐度的适应。泥盆纪已出现淡水双壳纲,在石炭纪、二叠纪时它们已形成一支相当大的淡水种族。到中、新生代它们更成为淡水环境的主要门类之一。其中一部分可能是古生代淡水种类的后裔(如费尔干蚌),而大部分则起源于海生种类(如蚬类、类三角蚌等)。它们首先从海水进入并适应淡化海水水体,如潟湖、三角洲等,然后经由河流到达湖泊、沼泽,很快地占领了底栖滤食的生态环境。但双壳纲毕竟不能像腹足纲那样登上陆地,这是受其内因(靠水流摄食和呼吸)制约的。
三、头足纲
(一)一般特征
头足纲(Cephalopoda)是软体动物门中发育最完善、最高级的类群,包括在地史时期曾极度繁盛并具有重要意义的角石、杆石、菊石、箭石和现代的鹦鹉螺、章鱼、乌贼等:它们全为海生的肉食性动物,善于在水底爬行或水中游泳。头足纲动物两侧对称. 头在前方而显著,头部两侧具有发达的眼,中央有口,口内有角质颚片。腕的一部分环列于口的周围,用于捕食;另一部分则靠近头部的腹侧. 构成排水漏斗,是独有的运动器官(图5-19)。它们的神经系统、循环系统和感觉器官等都较其他软体动物发达。雌雄异体。鳃有四个或两个,四鳃者具有外壳,如Nautilus(鹦鹉螺)(图5-19);二鳃者壳体被外套膜包裹而成内壳或无壳,如 Sepia(乌贼)、Octopus(章鱼)。
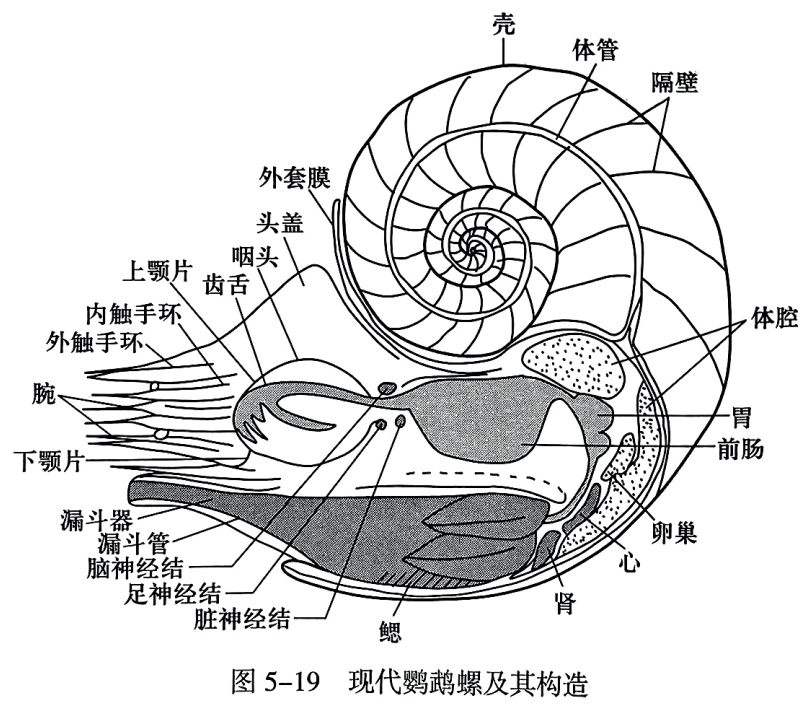
根据软体与硬体(壳体)的关系,可将头足纲动物分为两大类,即外壳类(ectoconchs)和内壳类(endoconchs),其中外壳类头足动物化石较丰富,在此重点介绍。
(二)头足纲外壳类动物壳的基本特征
1. 壳形
头足纲外壳类动物壳的形状多种多样,有直形、弓形、环形和旋卷形等(图5-20),少部分为锥形和螺旋形。壳体大小差别很大,一般壳的直径为数厘米或十余厘米,小的壳直径不到1cm,而最大的壳直径可达2.5m。比较而言,其中的菊石类壳的形状以旋卷形壳占绝大多数(图5-20之5—9)。鹦鹉螺类壳的形状则以简单的直锥形或弯弓形为主,也有旋卷的鹦鹉螺形和一些变异的环角石形和喇叭角石形等(图5-20之1—4)。在平旋壳形中,壳每旋转一周称为一旋环(whorl);最后旋成的旋环为外旋环(outer whorl);外旋环以内的所有旋环统称为内旋环(inner whorls)。根据旋卷程度,即相邻两个旋环之间包裹的程度可区分为外卷(evolute,仅接触在一起或包裹很小一部分)、半外卷(semi-evolute,包裹不超过侧面一半)、半内卷(semi-involute,包裹超过一半)和内卷(involute,外旋环完全包围内旋环)。
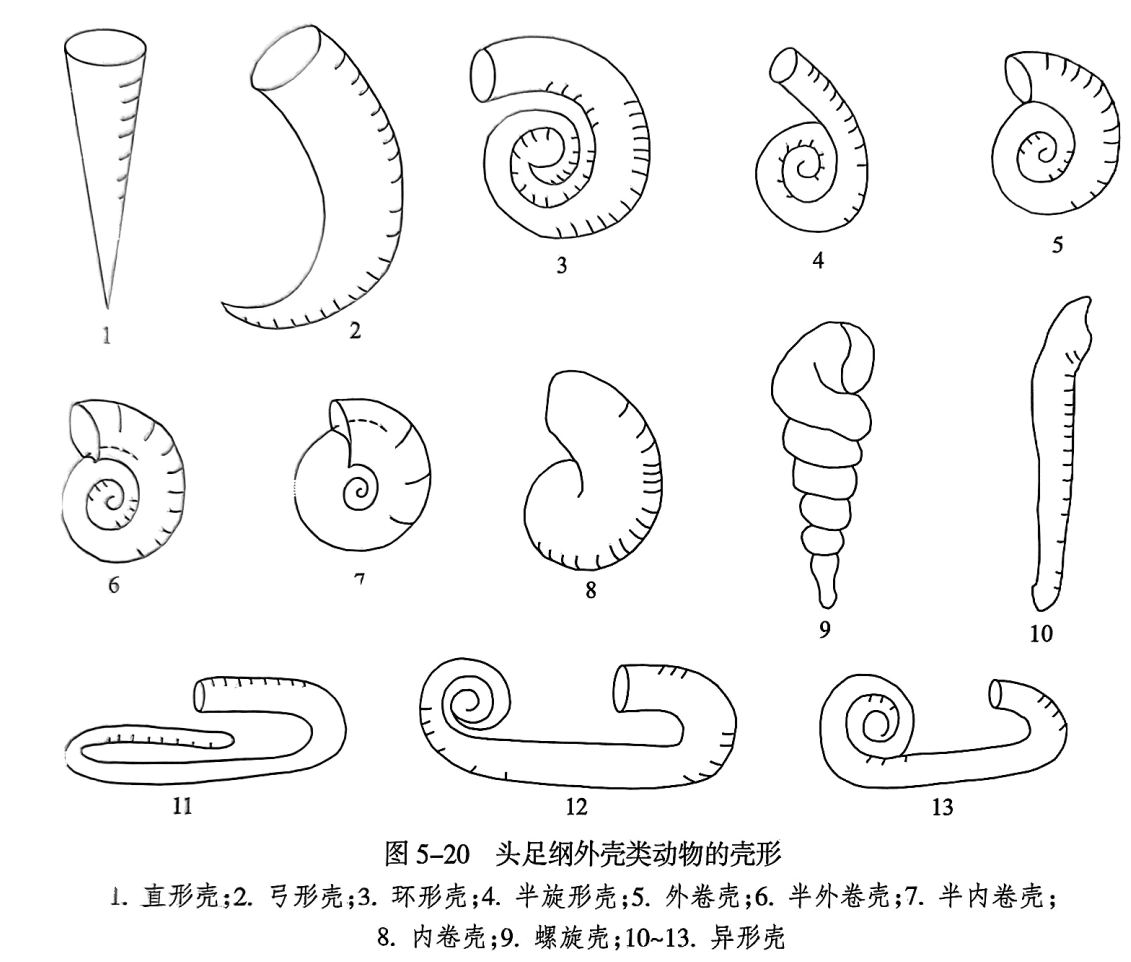
2. 壳的定向
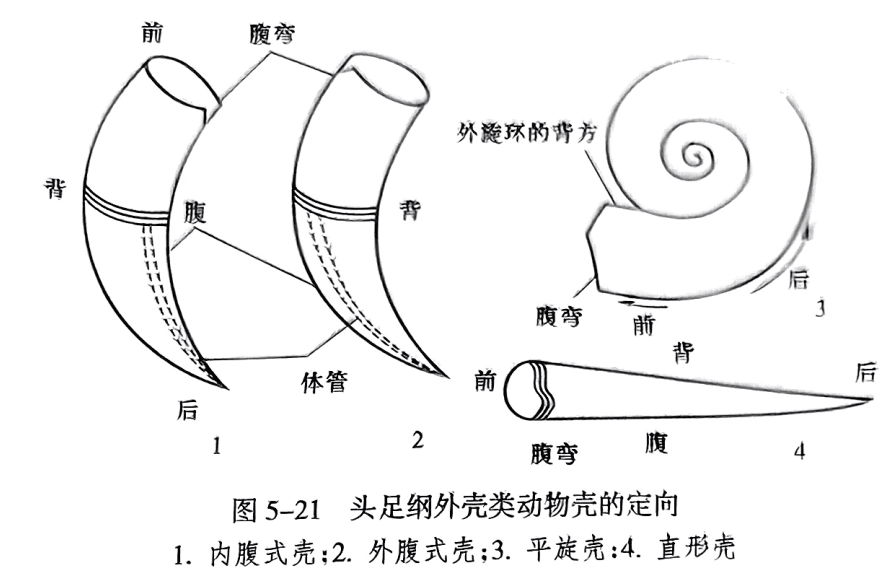
在直壳或弯壳中,壳的尖端为后方,壳的口部为前方;生长线通常向后弯曲;与体管靠近的一侧为腹方,另一方则称背方(图5-21)。在平旋壳中,壳口为前方,原壳为后方;旋环的外侧为腹方,内侧为背方(图5-21之3)。在弓形壳中,腹部位于凹侧者为内腹式壳(图5-21之1),反之为外腹式壳(图5-21之2)。
3. 壳的基本构造
壳体最初形成的部分为原壳(protoconch);随着生物体的增长和前移,软体周缘部分分泌其壳壁,而后缘部分分泌横向隔壁(septum,复数septa)来支持软体。随着生物体不断前移,外壳壁继续增长,而软体后部逐步和原来的隔壁分离;达到一定距离后,生物体前移暂时停止,又产生了新的隔壁;这样隔壁就将壳体分隔为许多房室(chamber);最前方具壳口的房室最大,为软体居住之处,叫住室(living chamber);其余各室充以气体称气室(air chamber)。平旋壳体的两侧中央下凹部分称为脐(umbilicus);脐的四周壳面叫脐壁(umbilical wall);脐壁外缘与旋环侧面相交处称为脐线或脐缘(umbilical margin);脐部相邻两旋环的接触线称脐接线(umbilical seam)(图5-22)。
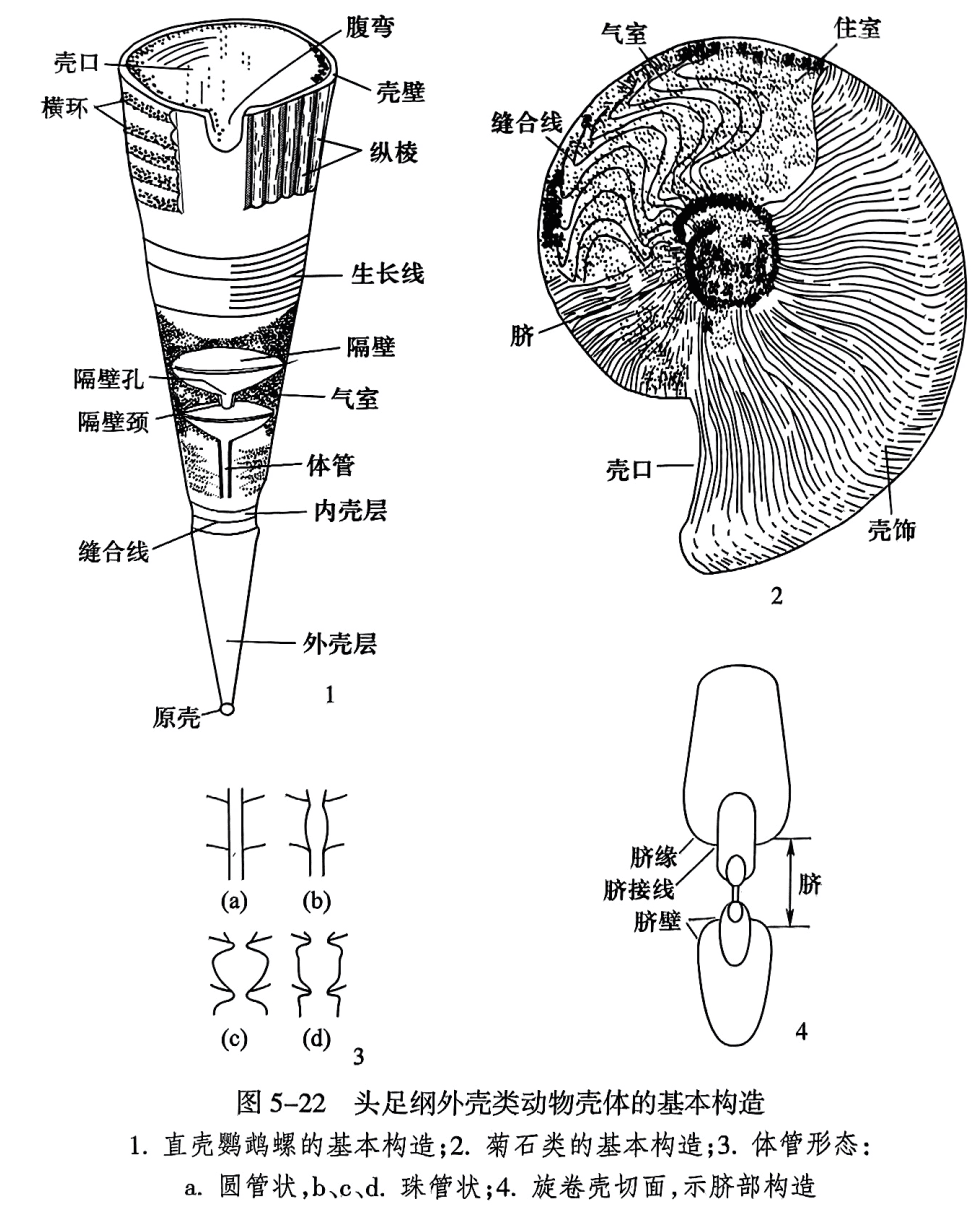
头足纲动物软体的后端生有一个肉质索状管(体管索,siphuncular cord),自住室穿过各气室而达原壳. 将软体拴在壳内。因此每个隔壁上都有被体管索所穿过的隔壁孔。沿隔壁孔的周围延伸出的领状小管称为隔壁颈(septal neck);隔壁颈向后方或外后方弯曲,其长度不一,短者1~2mm. 长者可达1~2个气室。隔壁颈之间或其内侧常有环状的钙质小管相连,这种环状物称为连接环(connecting ring)。连接环厚薄不一,简单或复杂。这些隔壁颈和连接环互相连接成为一条贯穿整个壳体的灰质管道,称为体管(siphuncle),它包围着软体的肉质体管索。体管一般位于壳体的中央或腹部,少数位于背部。体管的形状一般为细长的圆柱形,但也有些呈串珠形。
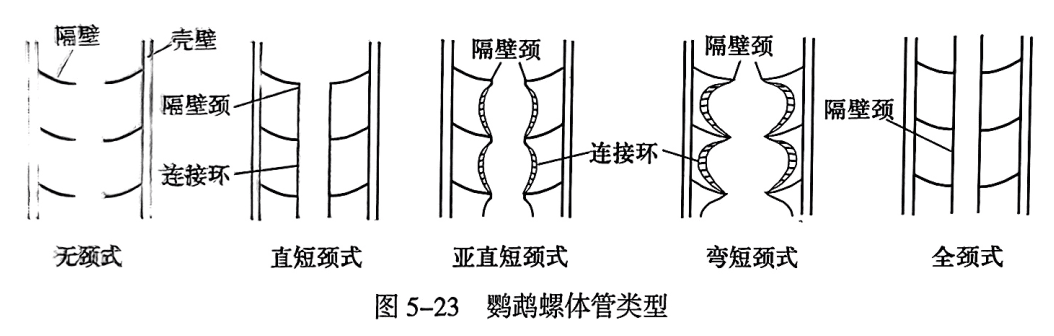
根据隔壁颈的长短、弯曲程度和连接环形状,可将体管区分为5种类型(图5-23):隔壁颈甚短或无、无连接环的无颈式(achoanitic);隔壁颈短而直、连接环直的直短颈式(orthochoanitic);隔壁颈短而直. 仅尖端微弯、连接环外凸的亚直短颈式(suborthochoanitic);隔壁颈短而弯、连接环外凸的弯短颈式(cyrtochoanitic);隔壁颈向后延伸,达到或超过后一隔壁、连接环或有或无的全颈式(holochoanitic)。
4. 壳饰
大多数鹦鹉螺壳的表面是光滑的或具有细的生长线,但有些类别的壳面具有横肋或纵纹,还有一些具有瘤或刺。菊石类壳的表面常发育与壳口边缘相平行的生长纹、生长线(图5-22之2). 与壳体旋卷方向一致的纵纹(旋纹),或与壳体旋卷方向相垂直的横肋。菊石类的横肋类型繁多,按其分布特征可分为疏的、密的和束状的;按其形状特征可分为凸形、凹形、双凹形、波形、镰形、之字形及闭锁形等。有时横向与纵向纹线相交形成网状饰。不少类别还具有壳刺和瘤状突起。某些种类还具有收缩沟构造,其方向与生长线相一致,在每一旋环上有3~10条不等。
5. 缝合线
头足纲动物壳内隔壁边缘与壳壁内面相接触的线称为缝合线(suture line),因此只有外壳表皮被剥去以后,才能露出缝合线(图5-22之2)。隔壁不褶皱的类别,其缝合线平直;反之缝合线显著地弯曲。平旋壳形的缝合线可以分为内外两部分,自腹中央经两侧面到脐接线的部分为外缝合线(external suture);自脐接线经过背部到另一侧的脐接线部分称为内缝合线(internal suture)。缝合线向前弯曲的部分称为鞍(saddle),向后弯曲的部分称为叶(lobe)。最基本的叶有4个(图5-24A):位于腹部的叶称为腹叶(ventral lobe),位于背部的叶称为背叶(dorsal lobe),位于侧面的叶称为侧叶(laterallobe),位于脐部的叶称为脐叶(umbilical lobe)。由脐叶或脐鞍再分出许多小的叶和鞍,合称为助线系(auxiliary)。在侧叶与脐叶之间若另有一侧叶,则前一侧叶称为第一侧叶(first lateral lobe),后者称为第二侧叶(second lateral lobe)。腹叶与第一侧叶之间的鞍称为第一侧鞍(first lateral saddle),第一侧鞍背方的鞍称为第二侧鞍(second lateral saddle)。有些类别由第一侧鞍再进行分化,形成了形似第一侧叶的次级叶,称为偶生叶(adventitious lobe)。内缝合线的叶和鞍大致和外缝合线相似. 除背叶外,亦有特征相似的侧叶和侧鞍,在描述时加“内”字,以区别外缝合线上的叶和鞍(图5-24A)。鹦鹉螺类的缝合线甚为简单,一般为环形。只有晚古生代的一些科属中缝合线发生弯曲,向前弯曲部分为鞍,向后弯曲部分为叶。
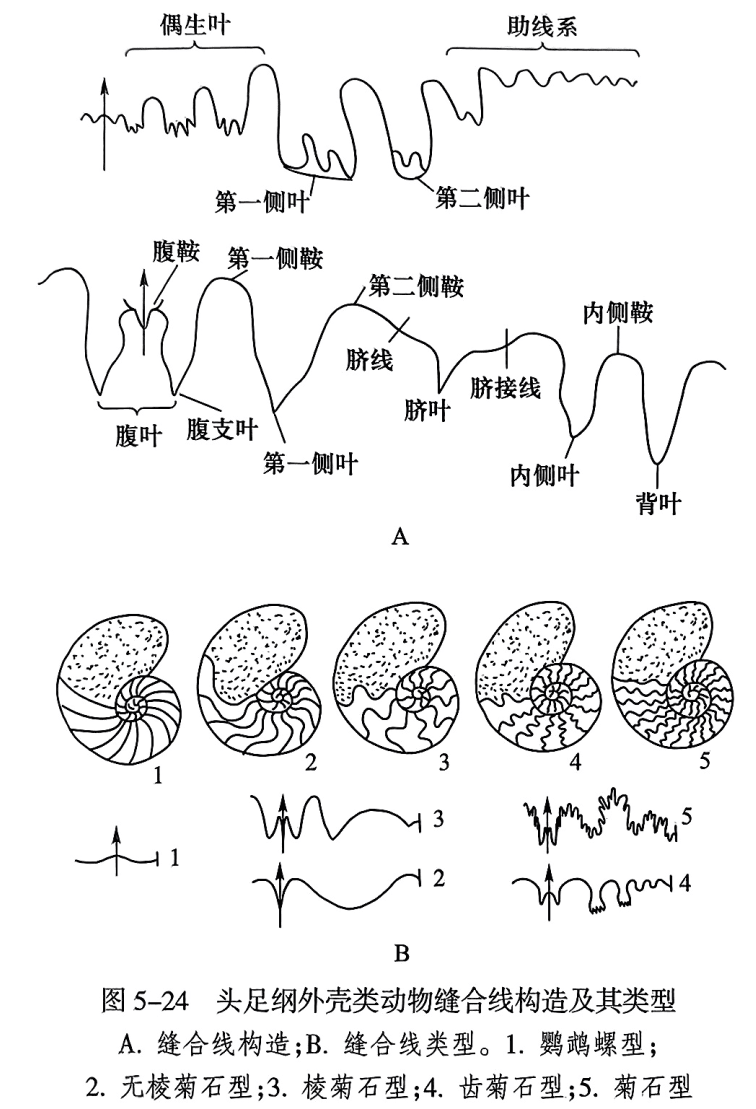
头足纲外壳类动物缝合线根据隔壁褶皱的程度,分为5种类型(图5-24B):
(1)鹦鹉螺型(nautiloid)。缝合线甚为简单,一般为环形,没有明显的鞍叶之分。
(2) 无棱菊石型(agoniatitic)。鞍叶数目少,形态完整,侧叶宽,浑圆状。
(3)棱菊石型(goniatitic)。鞍叶数目较多,形态完整,叶常呈尖棱状。
(4)齿菊石型(ceratitic)。鞍部完整圆滑,叶部再分为齿状。
(5) 菊石型(ammonitic)。鞍和叶再分出许多小叶。
(三)头足纲的分类
古生物学界对头足纲的分类意见分歧较大,提高和增加分类级别是目前分类工作的主要倾向。目前一般将头足纲动物分为4个亚纲:鹦鹉螺亚纲(Nautiloidea)、杆石亚纲(Bactritoidea)、菊石亚纲(Ammonoidea)和鞘形亚纲(Coleoidea),前3个亚纲为外壳类,鞘形亚纲为内壳类。表5-6列举了头足纲动物外壳类各亚纲的主要特征。

(四)头足纲动物的生态与形态功能分析
现代的头足纲动物都是海生的,它的化石都保存在含有其他各种海生生物化石的地层中,因此可以认为地史时期的头足纲动物也都是海生的。
现代的内壳类头足纲动物,都是营游泳生活的,有的善于游泳,有的不善于游泳,而在海底爬行。中生代最典型的内壳类箭石由于有笨重的鞘,所以虽然能游泳,但其游泳能力较弱。
现代的外壳类头足纲动物仅存鹦鹉螺(Nautilus)一属,主要生活在太平洋南部、亚洲东部与澳大利亚之间的东印度群岛周围的浅海区,也可以到达较深的海区,营游泳及底栖爬行生活。从保存的外壳类头足纲动物化石来看,它们都具有气室,壳壁较薄,壳面的脊和瘤内部也是空的,说明其密度比较小,便于动物在水中游泳,因此人们推测外壳类头足纲动物都具有一定的游泳能力。由于它们具有各种不同的壳形,游泳能力有所差别。比较光滑扁平、特别是具有流线型的壳体的,是一种善于快速或长距离游泳的类型。两侧对称平旋状壳体,便于在水中保持平衡,显然是有利于游泳的壳形。在直锥形壳体中,长锥形壳,体管细小,气室内无沉积物的类型,一般是沿水平方向或尖端向上斜的游泳种类;而体管粗并靠近腹部,气室、体管沉积物发育者是栖息海底的类型。壳短粗呈流线型者,如袋角石类,它们在成年期时并不保存它整个发育期间的壳,表明其周期性地截去其壳的始端,即将早期形成的气室从壳体断去,变得短粗以减小始端浮力,而使壳体保持平衡。同时此类动物壳内无沉积物或沉积物很少,因此是善于游泳的动物。另一类壳体短粗,口部收缩为裂缝状的头足纲动物,是底栖爬行或浮游的类型。少数外壳类呈螺塔状或蠕卷状,由于壳体笨重,仅能在海底爬行或营底栖固着生活(图5-25)。
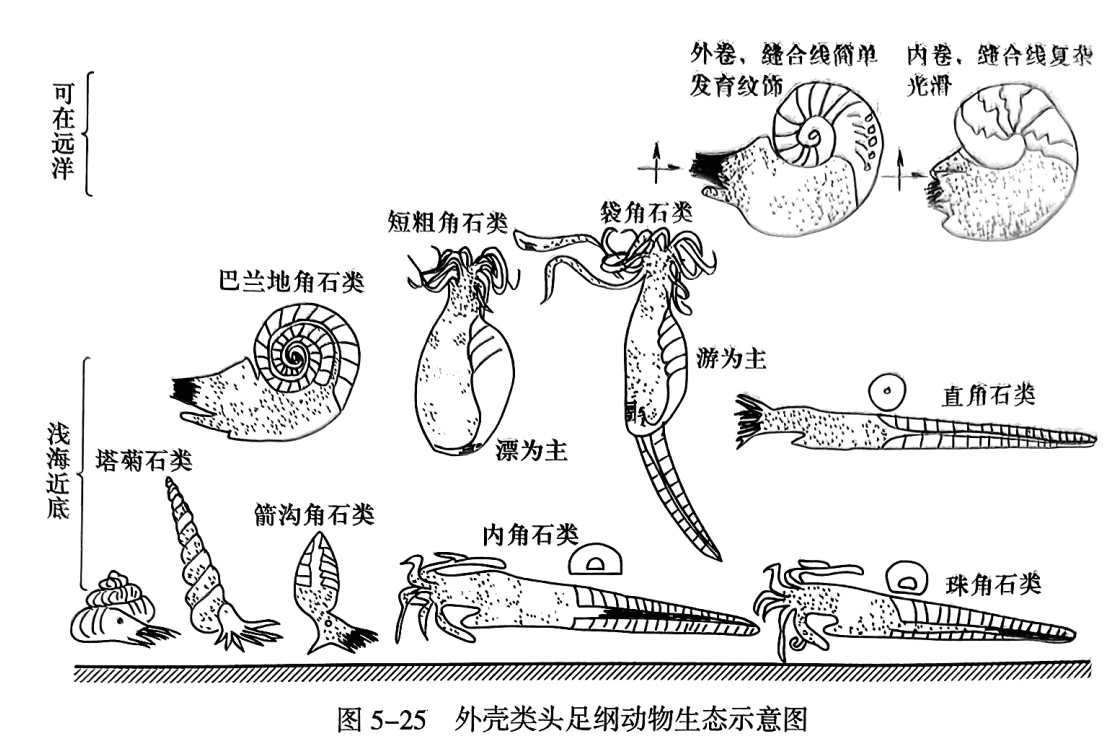
头足纲动物一般都有很广的地理分布,这一方面是由于大多数营游泳或漂游生活,可漂游至很远。另一方面,由于头足纲动物死后,软体腐烂或从外壳上脱落,这时充满气体的外壳便漂浮在水中,而被洋流运到远处。由于头足纲动物演化得很快,属种生存的地质时间短,再加上地理分布广的特点,使它们当中的许多种类成为标准化石,可以进行远距离地层的对比。
(五)头足纲动物的地史分布
鹦鹉螺类化石最早见于晚寒武世地层中,是一些形体小、构造简单、隔壁颈短和壳体略弯的类型。在奥陶纪时迅速发展,趋于极盛,壳体增大,不少属种的体管构造比较复杂,除直壳类型外还出现了平旋壳类型。至志留纪——泥盆纪时鹦鹉螺类开始衰落。在三叠纪末期,直壳类绝迹,旋卷类亦变少,至今只留下一属(鹦鹉螺属,Nautilus)。
菊石亚纲最早出现于泥盆纪初期,繁盛于中生代,在白垩纪末期全部灭绝。自泥盆纪至白垩纪末期,菊石亚纲有4个繁盛期:①晚泥盆世,由海神石类及无棱菊石、棱菊石类的低级代表组成;②晚石炭世至二叠纪末,主要是棱菊石类,其次为无棱菊石类的高级代表;③三叠纪,由齿菊石类组成;④早侏罗世至晚白垩世,由菊石类组成。
纵观头足纲动物的兴衰历史,大致可以分为3个阶段:①早古生代阶段,这一时期的头足纲动物全为鹦鹉螺类,其缝合线均为简单的鹦鹉螺型,在头足纲动物的演化史中处于早期阶段。②晚古生代至中生代阶段,这一时期以菊石类和箭石类为主,鹦鹉螺类居于次要地位。自泥盆纪开始,头足纲动物向高级方向发展,主要表现在:一是缝合线趋向复杂化,导致了早泥盆世低等菊石类的兴起;另一是由外壳转为内壳,衍生出早石炭世的箭石。菊石类和箭石类的兴起,是头足纲动物演化史上重要的转折点,由此进入了一个新的阶段。③新生代阶段,以内壳类的繁荣为特征,鹦鹉螺类仅个别种类残存于海洋中,箭石类亦在本时期初灭绝,鞘形亚纲的八腕类及除箭石类以外的十腕类十分繁盛,代表了头足纲动物发展史上的高级阶段。