7.5 爬行纲
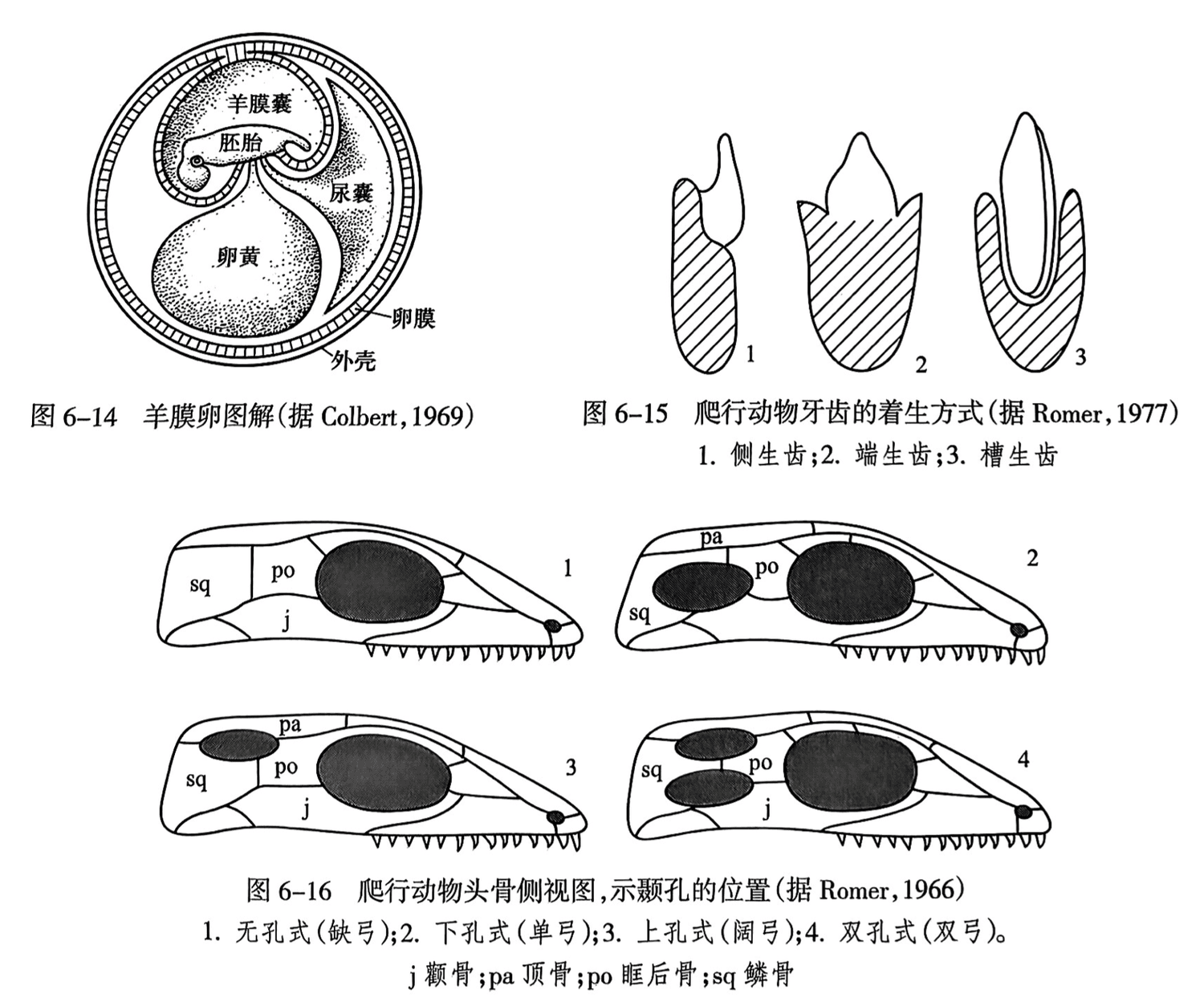
爬行纲(Reptilia),也称为爬行类(reptiles),最早出现于晚石炭世,从两栖动物的石炭蜥类祖先演化而来。爬行纲和两栖纲之间的本质区别是卵的结构不同:两栖纲的卵像鱼类的卵一样,无羊膜结构,必须产在水中,在水中孵化;爬行纲的卵有羊膜结构,故称羊膜卵(amnioticegg)(图6-14),可以产在陆地上和在陆地上孵化。爬行纲卵结构的这一进化特征使脊椎动物彻底摆脱了对水体的依赖,能在陆地上生活和繁殖后代。爬行纲及其衍生的后裔哺乳纲和鸟纲也因此总称为羊膜动物(amniotes)。
爬行纲的其他典型特征有:①头骨高,不同于两栖纲那种通常扁平化的头骨;②脊椎骨大部或完全由侧椎体构成;③原始的爬行纲有两块荐椎骨,不同于两栖纲只有一块荐椎骨,而进步的爬行纲可有多达八块的荐椎骨;④肩胛骨和乌喙骨显著加强,匙骨、锁骨和间锁骨缩小或消失;⑤体外被覆角质的鳞甲。
爬行纲的牙齿一般都是形态相似的圆锥形,称为同型齿(homodont)。牙齿着生于颌骨上的方式有三种:①侧生齿(pleurodont),牙齿基部附于颌骨内缘,血管与神经由侧面伸入(图6-15之1);②端生齿(arcodont),由纤维膜附于颌骨的边缘,无齿根,易脱落(图6-15之2);③槽生齿(thecodont),齿根发达,深植于颌骨的齿槽中,鳄类有此种牙齿,其与哺乳动物的牙齿性质相同(图6-15之3)。
爬行纲主要依据头骨颞孔的有无、数目,以及位置等分为如下四个亚纲:
一、缺弓亚纲或无孔亚纲(Anapsida)
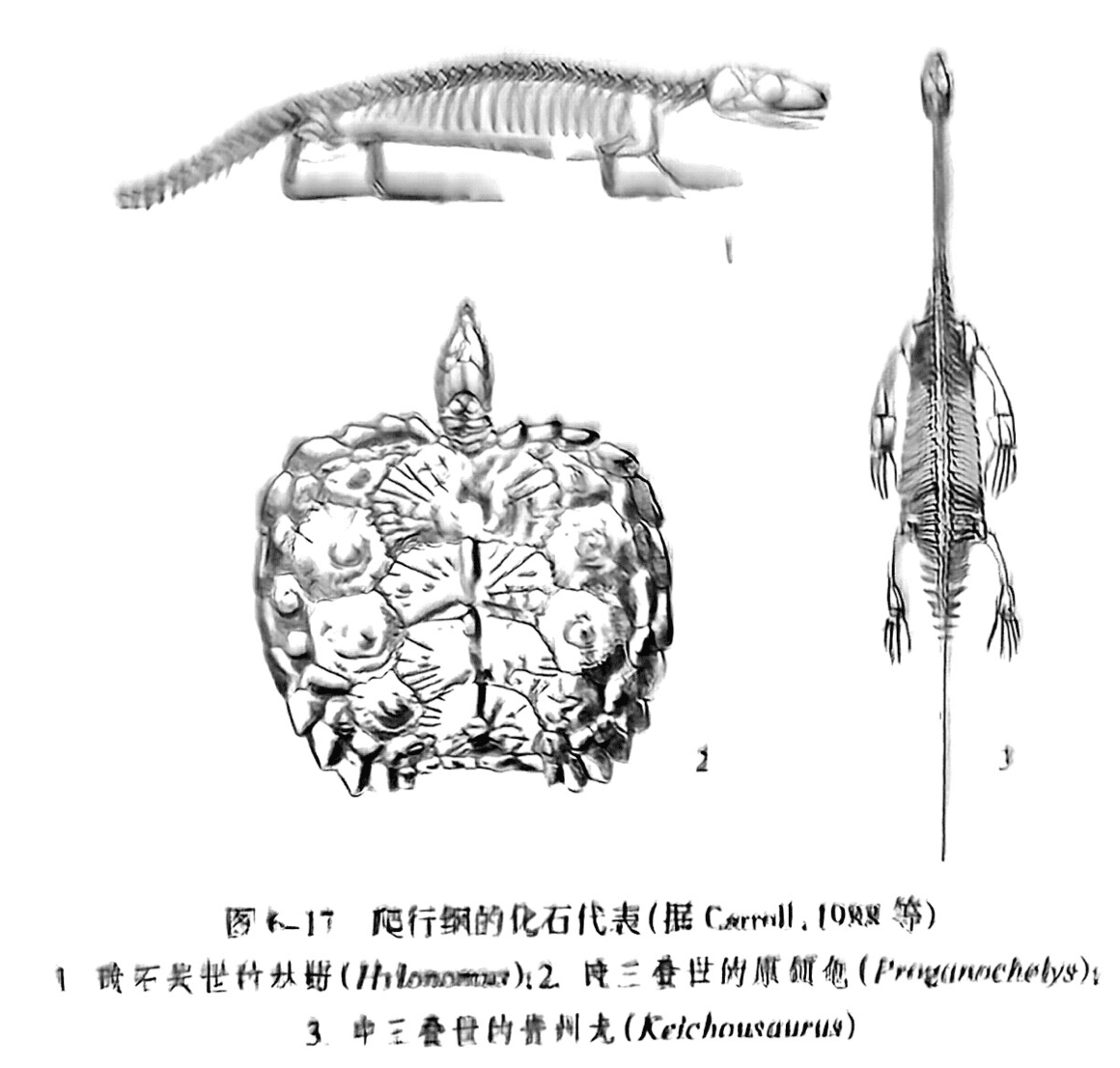
本亚纲见于晚石炭世至晚三叠世,无颞孔(图6-16之1)。本亚纲分为两个目:大鼻龙目(Captorhinida)和中龙目(Meseanria)。大鼻龙目是最原始的爬行类,几乎呈全球性分布,从中衍生出其他3个亚纲。在加拿大东部晚石炭世早期地层中发现的林蜥(Hylonomus)是大鼻龙目的早期代表,体长不超过1m(图6-17之1);我国大鼻龙目的代表有华北上二叠统孙家沟组中发现的石千峰龙(Shihtienfenia)。中龙目仅发现于南非和巴西的早二叠世陆相地层中,曾被魏格纳作为大陆漂移学说的重要证据之一。
二、龟鳖亚纲(Testudinata)
本亚纲见于晚三叠世至今,呈全球性分布。具有甲壳的爬行类,无颞孔,曾归入缺弓亚纲。由于其解剖学特征和生活习性跟大鼻龙目存在显著差别,所以本亚纲目前被作为一个独立的亚纲。最早的龟鳖类化石发现于德国和中国。德国上三叠统的原颌龟(Proganochelys)(图6-17之2).其头骨数目减少,颌骨边缘没有牙齿,躯体为厚重的壳所保护等特征显然已很像现存的龟鳖类。中国贵州晚三叠世地层也发现了完整保存的半甲齿龟(Odontochelys semitestacea)和中国始喙龟(Eorhynchochelys sinensis)。半甲齿龟口中还有牙齿,腹甲完整,背甲不完整;而中国始喙龟则是已知最早有喙的龟类。龟鳖亚纲在侏罗纪时便有了很大的发展,虽然在中生代末期大量的爬行动物相继灭绝,但是龟鳖亚纲依靠甲壳的保护繁衍至今。我国龟鳖亚纲的化石非常丰富,地理分布遍及全国各个省级行政区,时代从晚三叠世至今。
三、双弓亚纲或双孔亚纲(Diapsida)
本亚纲见于晚石炭世至今,呈全球性分布。本亚纲具有两个颞孔(图6-16之4)。除龟鳖亚纲之外的所有现代爬行动物都隶属该亚纲。鳄、蜥蜴、蛇和喙头蜥的祖先可以追溯到中生代早期、双弓亚纲是爬行纲中多样性最高的门类,包括中生代的陆上霸主恐龙、展翼翱翔的翼龙,以及重返海洋的蛇颈龙、鱼龙等,因此对它们的研究也最深入。本亚纲分为鱼龙形次亚纲、鳞龙形次亚纲、初龙形次亚纲和海龙形次亚纲。
1 .鱼龙形次亚纲(Ichthyosauromorpha)
本次亚纲见于早三叠世至晚白垩世,包括湖北鳄类(hupehsuchians)和鱼龙形类(ichthyusauriforms),几乎呈全球性分布。鱼龙形类是高度特化的海生爬行动物,只有一个上颞孔。由后额骨和上颞骨构成其上界(图6-16之3)。有类似鱼的背鳍、胸鳍、腹鳍和尾鳍,脊柱伸入尾下叶;头大,眼大,吻长,有长的利齿,肉食。我国的鱼龙形次亚纲化石集中于三叠纪,分布地区颜广,在西藏、贵州、云南、安徽、湖北均有发现。
2. 鳞龙形次亚纲(Lepidosauromorpha)
本次亚纲的四肢像原始的四足动物那样侧向伸展,保持爬行的运动姿势。脊柱的侧向扭动成为运动的一个动力,在蛇类中表现得最为明显。本次亚纲包括始鳄目(Eosuchia)、喙头目(Rhynchocephalia)、有鳞目(Squamata)、楯齿龙目(Placodontia)、幻龙目(Nothosuuria)和蛇颈龙目(Plesiosauria)。
广义的始鳄目包括双弓亚纲的基干类群,从晚石炭世延续到始新世。在南非上二叠统中发现的小而原始的似蜥蜴的杨氏蜥(Youngina)被认为是现代有鳞目和喙头目的祖先。喙头目从早三叠世延续至今。云南禄丰和辽宁凌源上三叠统中的禄丰喙头蜥(Lufengocephalus)和喙头龙(Rhynchosaurus)等均为该类的代表。有鳞目从晚二叠世延续至今,是现代爬行纲中的常见者,代表性的种如蜥蜴、蛇。我国有鳞目化石亦不少,如晚侏罗世的矢部龙(Yabeinosaurus)等。有鳞目和喙头目合称鳞龙超目(Lepidosauria)。
楯齿龙目、幻龙目和蛇颈龙目合称蜥鳍超目(Sauropterygia)。楯齿龙目是蜥鳍超目其他两个目的姐妹群,生活的时代仅限于三叠纪,分布于东、西特提斯区,在我国的代表有贵州中-上三叠统的中国豆齿龙(Sinocyamodus)。幻龙目包括肿肋龙科(Pachypleurosauridae)和幻龙科(Nothosauridae).生活的时代也限于三叠纪,是仅发现于欧洲和我国的一类已灭绝的水生爬行动物。在贵州发现的贵州龙(Keichousaurus)是肿肋龙科的代表(图6-17之3),而欧龙(Lariosaurus)是幻龙目的代表。蛇颈龙目生活的时代从早侏罗世延续到晚白垩世,中国上龙(Sinopliosaurus)是其代表。
3. 初龙形次亚纲(Archosauromorpha)
本次亚纲脊柱的侧向扭动已大大减弱,四肢不侧向伸展,成行走姿势。本次亚纲包括八个目,其中原龙目(Protorosauria)、三棱龙目(Trilophosauria)和喙龙目(Rhynchosauria)为该次亚纲的基干类群,其余五个目为进步的双孔类,合称初龙超目(Archosauria)。现简述如下:
(1)槽齿目(Thecodontia)。本目生活于晚二叠世至晚三叠世,是原始的初龙类。我国山西、新疆上三叠统中有许多槽齿目化石。
(2)鳄目(Crocodilia)。本目生活于晚三叠世至今,由槽齿目演化而来。在中生代时相当繁盛,在新生代时逐渐衰落,现存仅20余种,包括产于长江中下游的扬子鳄。
(3)翼龙目(Pterosauria)。本目生活于晚三叠世至晚白垩世,是恐龙类的近亲。其特征是前肢第四指骨特别加长支撑由身体侧面延展的皮膜,变成翅膀,适于空中飞行,胸骨也相应地变得发达,很可能是温血动物。早白垩世的准噶尔翼龙(Dsungaripterus)是我国发现的第一件完整的翼龙化石,在辽西热河生物群地层中也发现了很多翼龙目化石,其中主要是较进步的翼手龙类。
(4)恐龙(Dinosauria)。恐龙是初龙形次亚纲中的一个单系类群。1841年,英国学者欧文发现了一些体型巨大的爬行动物化石,其腰带和股骨的形态表明它们能够直立行走,于是他首次提出了“恐龙”(dinosaur)一词。按照分支系统学的定义,恐龙是指三角龙与现代鸟类的最近共同祖先及其所有后裔。这一类群的主要鉴定特征包括后额骨缺失、肱骨三角胸嵴发育、肠骨后支腹缘具有一短窝、髋臼穿孔、胫骨远端横向扩展呈近长方形,以及距骨前缘有一个上升支。恐龙可以划分为蜥臀目(Saurischia)和鸟臀目(Ornithischia)。目前最早的恐龙骨骼化石来自于晚三叠世卡尼期(约2.3亿年前)地层,其中化石最丰富的地方为阿根廷和巴西。另外足迹化石表明恐龙可能出现得更早,在中三叠世甚至早三叠世就已经出现了恐龙的直系祖先类群。恐龙繁盛于侏罗纪和白垩纪。我们通常所称的恐龙(或非鸟恐龙)灭绝于6600万年前的白垩纪末期。世界上迄今已命名的恐龙有1000多种。
不同的恐龙个体差别很大,体长从不足一米到几十米,生活方式也不尽相同:有些营群居生活,有些则单独生存,有些是植食者,有些则是肉食者。蜥臀目和鸟臀目之间的主要区别在于腰带构造上的不同(图6-18之1)。蜥臀目的腰带形态较接近蜥蜴,其耻骨仅向下前方一个方向延伸,从侧面观察呈三射式。鸟臀目的耻骨向前有一个突起,向后延伸与坐骨平行,从侧面观察腰带呈四射式结构,与鸟类的腰带结构相似。
蜥臀目根据动物行走运动的姿态分为两个亚目,即兽脚亚目(Therproda)和蜥脚亚目(Saurpoda),
兽脚亚目包括所有双脚行走的蜥臀类。白垩纪的霸王龙(Tyrannosdurus)是该亚目最著名的代表.这是一种大型的食肉恐龙,长12~14m,后脚粗壮,前脚缩小,头骨巨大,具有刺刀状的利齿.颈短(图6-18之3)。在中国东北部的燕辽生物群(中、晚侏罗世)和热河生物群(早白垩世)地层中发现的大量有羽毛的恐龙,如近鸟龙(Anchiornis)、小盗龙(Microraptor)、中华龙鸟(Sinosauropteryx)、尾羽龙(Caudipteryx),为论证鸟类起源于小型兽脚类恐龙的假说,特别是羽毛的演化和鸟类飞行的起源提供了大量的证据(图6-18之2)。
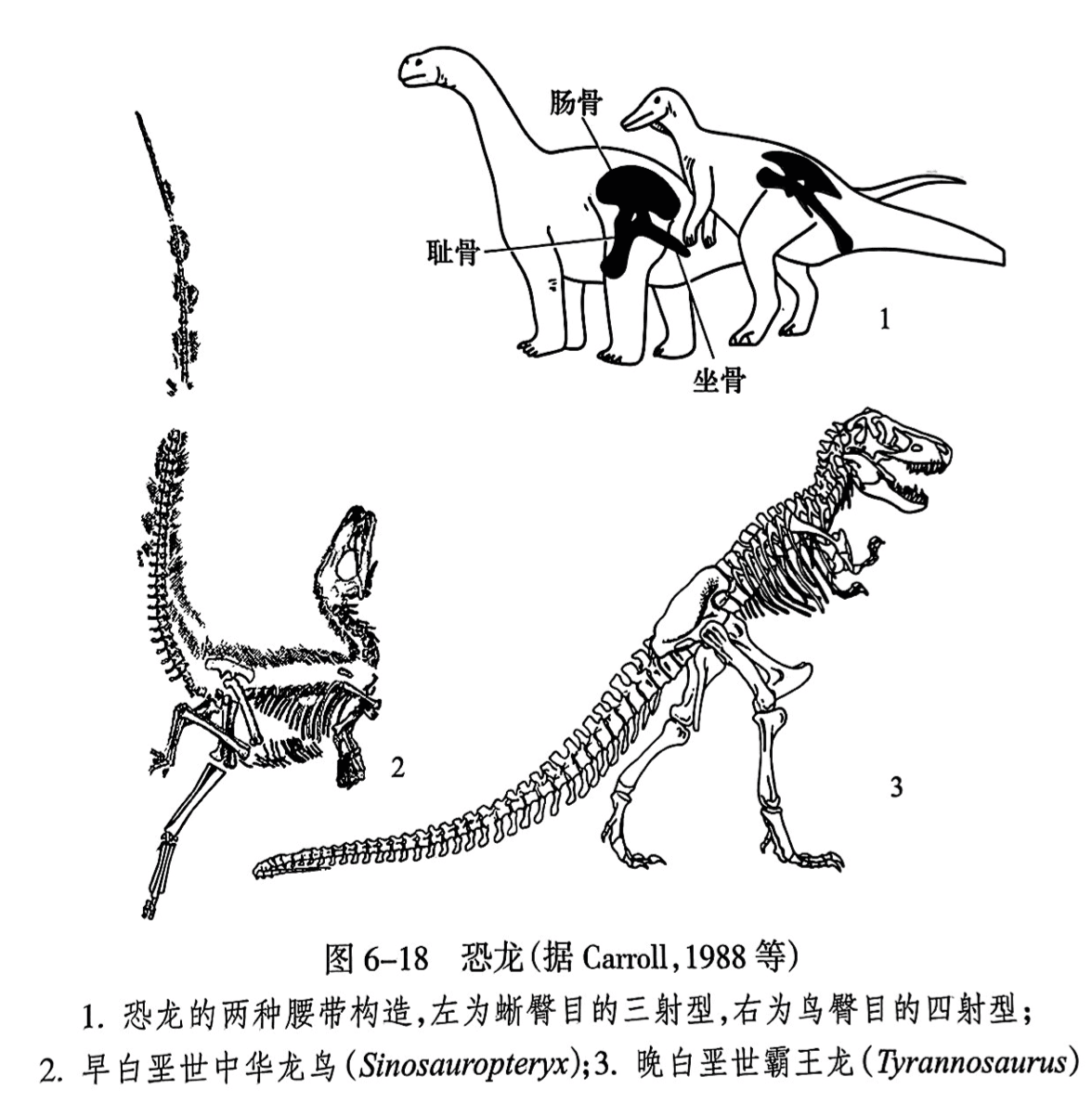
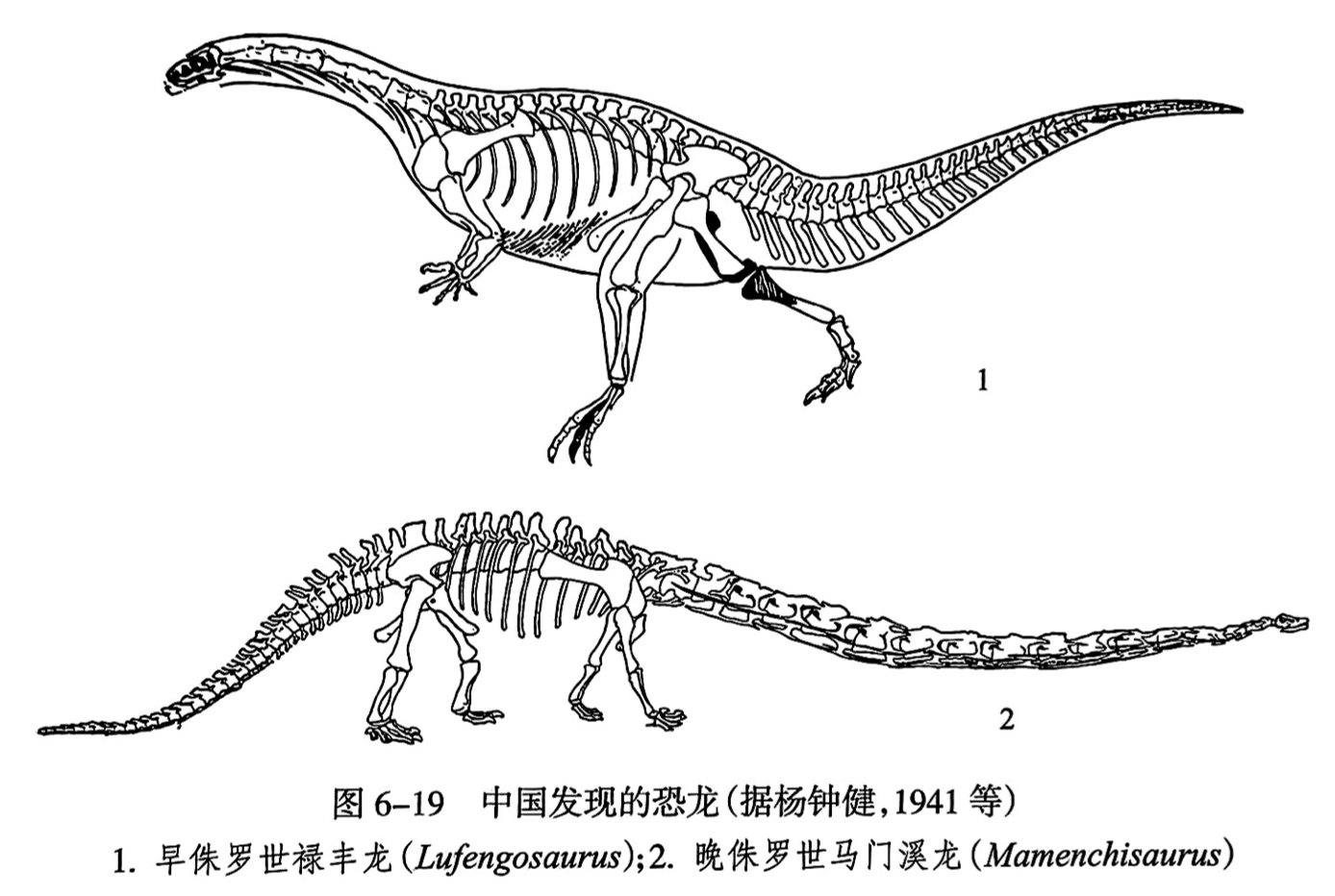
蜥脚亚目是四足行走的蜥臀类。我国云南禄丰早侏罗世发现的禄丰龙(Lufengosaurus)和云南龙(Yunnanosaurus)是原始蜥脚亚目的重要代表,仍为两足行走(图6-19之1)。而进步的蜥脚亚目是真正四足行走的,其特征是身体巨大,其包括世界上已知最大的恐龙,长可达35~40m,重达80吨以上,头很小,颈长,尾长。我国在四川盆地的合川、宜宾和甘肃永登发现的马门溪龙(Mamenchisaurus)就属于此类(图6-19之2)。
鸟臀目包括原始的异齿龙科(Heterodontosauridae)及五个亚目,即鸟脚亚目、剑龙亚目、甲龙亚目、角龙亚目和肿头龙亚目。异齿龙科的典型代表为南非旱侏罗世地层中发现的异齿龙(Heterodontosaurus),其吻部具有大的犬齿。鸟脚亚目包括一些原始的小型类群和一些进步而体型较大的禽龙类(Iguanodontia)和鸭嘴龙类(Hadrosauroidea),如在山东莱阳上白垩统中发现的青岛龙(Tsintaosaurus),其头顶上长有奇特的棘突。角龙亚目的化石代表是热河生物群的常见 类别———鹦鹉嘴龙(Psittacosaurus),以及北美洲晚白垩世地层中发现的三角龙(Triceratops)等。肿头龙亚目因头骨肿大成圆顶状或平顶而得名,这种结构可能用于抵御捕食者或者同类竞争,它们主要生活在晚白垩世亚洲和北美洲。剑龙亚目背部有两排竖直的骨板,尾部有成对的、尖锐的尾刺,如四川自贡晚侏罗世的沱江龙(Tuojiangosaurus)。甲龙亚目也是体表被覆骨片的一类中等大小的植食性恐龙,有的尾部具有粗壮的尾锤。
4.海龙形次亚纲
海龙形次亚纲(Thalattosauromorpha)是三叠纪海生爬行动物中的一个单系类群,为初龙形次亚纲和鳞龙形次亚纲的姐妹群。海龙形次亚纲的体长可达3m以上,头后骨骼仍然保留了陆生四足类的特征,几乎没有什么适应水生生活的改变。贵州中-上三叠统中的安顺龙(Anshunsaurus)是在我国首次发现的海龙形次亚纲化石。
四、单弓亚纲或下孔亚纲(Synapsida)
本亚纲生活于晚石炭世至晚三叠世,具有一个颞孔,由眶后骨和鳞骨构成其上界(图6-16之2)。它们是联接原始爬行动物和哺乳动物的桥梁,早期成员与大鼻龙目十分接近,晚期成员则发育了一些类似哺乳动物的特征,如牙齿分异,肩带有前乌喙骨和乌喙骨等,因此本亚纲又称为似哺乳爬行动物。本亚纲的进步成员与哺乳动物很难划分,根据目前的观点,哺乳动物定义为下颌只有一块齿骨,否则便不能归入哺乳动物之中。本亚纲分为两个目:盘龙目和兽孔目。
(1)盘龙目(Pelycosauria)。本目为原始的似哺乳爬行动物,生活于晚石炭世至晚二叠世,分布于北美、欧洲和非洲。
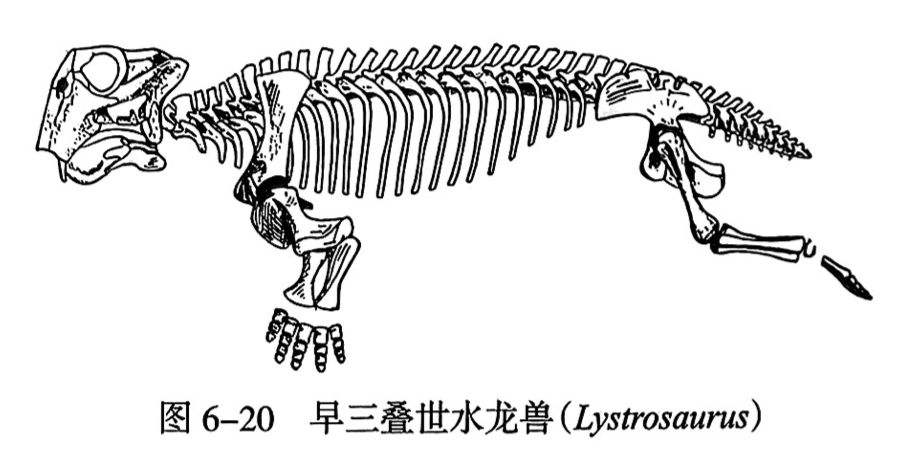
(2)兽孔目(Therapsida)。本目为进步的似哺乳爬行动物,生活于晚二叠世至早侏罗世,呈全球性分布,中国和南非是此类化石的著名产地。它们的颞孔十分宽大,进步的兽孔目的颞孔上缘不是由眶后骨和鳞骨构成的,而是由顶骨构成的,方骨与方轭骨则缩小。齿骨增大,牙齿分化为门齿、犬齿和颊齿。俄罗斯晚二叠世的渐衰鳄(Phthinosuchus)是该目的原始代表。由渐衰鳄开始.兽孔目沿着两条大的辐射适应路线向前发展。一支是异齿兽类(anomodonts).其中包括大型的恐头兽类(dinocephalians)和分布广泛的二齿兽类(dicynodonts)。三叠纪的水龙兽(Lystrosaurus)(图6-20)和中国肯氏兽(Sinokannemeyeria)都是二齿兽类的重要代表。另一支是最终衍生出哺乳动物的兽齿类(theriodonts),云南禄丰早侏罗世产出的卞氏兽(Bientherium)是其著名的代表。