8.5裸子植物
裸子植物(gymnosperms)是介于蕨类植物和被子植物之间的一类维管植物,它和苔藓、蕨类植物最大的不同点是能产生种子,但种子裸露,没有被果皮包被。裸子植物大多数为乔木,有强大的根系.维管系统发达.具有形成层,次生生长,包括种子蕨植物门、苏铁植物门、银杏植物门、松柏植物门、买麻藤植物门等。此外,还有一类介于蕨类植物与裸子植物之间的维管植物——前裸子植物门。本书也归到裸子植物中来介绍。
一、前裸子植物门(Progymnospermophyta)
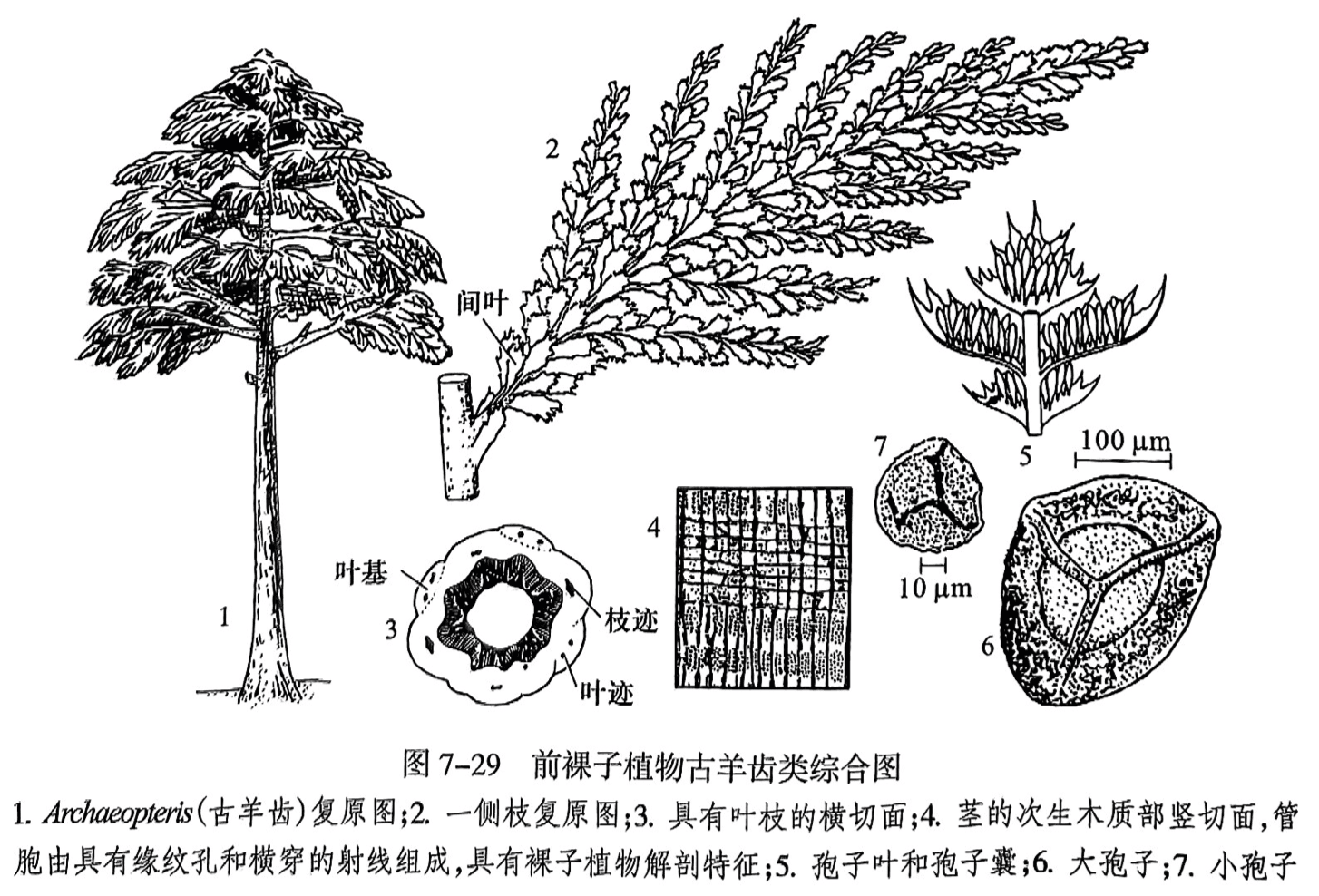
仅在泥盆纪至二叠纪早期地层中发现,其茎干的次生木质部为具有缘纹孔的管胞组成(图7-29之4),这是裸子植物茎的典型解剖特征;但生殖器官为蕨类植物型,即孢子囊(图7-29之6).因而视为蕨类植物和裸子植物之间的过渡类群,也有很多学者认为它是裸子植物的祖先类型而置于裸子植物中。本植物门以古羊齿类为最典型代表。近些年在我国内蒙古乌海发现整个树冠得以保存的瓢叶目化石 Paratingia wuhai(乌海拟齿叶),也较好地揭示这类植物具有裸子植物木材结构和孢子植物繁殖方式的特点。
二、种子蕨植物门(Pteridospermophyta)
1.一般特征
种子蕨植物门是古老的裸子植物,化石大多数保存为大型羽状复叶,叶的形态与真蕨植物门的叶几乎无法区别,但生殖叶上长有种子(图7-30之1),故取名为种子蕨。植物体为小乔木或灌木,也有藤木,有些很少分枝,这与苏铁植物相似。

2.蕨形叶形态分类
种子蕨和真蕨叶形十分类似,在古植物学中统称为蕨形叶(fernlike foliage)。由于化石保存原因,古生代地层中的很多蕨形叶化石缺乏生殖器官,无法确定其自然分类位置,因此常采用形态分类的办法,即依据蕨形叶的形状、脉序型式、小叶与轴的关系等特征建立形态分类及形态属。蕨形叶分为很多种形态类型,常见的有:①扇羊齿型(小羽片菱形、楔形,扇状脉);②楔羊齿型(小羽片边缘分裂,基部收缩,纤弱羽状脉);③栉羊齿型(小羽片椭圆形,基部全部附着于轴,羽状脉);④脉羊齿型(小羽片基部收缩成心形,以一点附着于轴,羽状脉,中脉不达顶端即消散);⑤座延羊齿型(小羽片基部下延,具有邻脉);⑥带羊齿型(带状叶或带状小羽片,中脉粗直,侧脉与中脉夹角较大)。在此基础之上进行蕨形叶形态分类,具体结果如表7-3和图7-31。
3.地史分布及生活型
种于蕨植物始现于晚泥盆世,在石炭纪至中二叠世极盛,在晚二叠世衰退,在中生代发展出新类型.但在早白垩世后全部灭绝。种子蕨植物的生活型多种多样,大多数是灌木状藤本或藤本植物。具有细长的茎和大的羽状复叶或单叶;有的如现代的树蕨一样,具有高而分枝少的茎和大型羽状复叶:古生代—中生代之交的舌羊齿具有粗壮主茎和大的单叶。

三、苏铁植物门(Cycadophyta)
1.一般特征
现代苏铁植物多数是茎粗矮的常绿木本植物,而化石苏铁常为细茎类型;很少分枝或不分枝;茎顶端丛生坚硬革质的一次羽状复叶或单叶,羽状分裂的裂片着生于羽轴两侧或羽轴腹面;大多数具有平行脉、放射脉,个别为单脉或网状脉;叶表皮角质层厚,气孔下陷;生殖器官聚集成球花(孢子叶球),位于茎的顶端;雌雄异株或同株;茎的解剖结果显示皮层和髓部发育,而次生木质部很薄。
2.分类
由于苏铁植物在地层中经常见到的是叶的印痕化石,所以它们单凭叶的外部形态不易进行自然分类,因而常常根据叶形态先建立形态属,再根据表皮细胞结构进行自然分类。苏铁植物依据生殖器官和叶的解剖特征分为苏铁纲和本内苏铁纲。
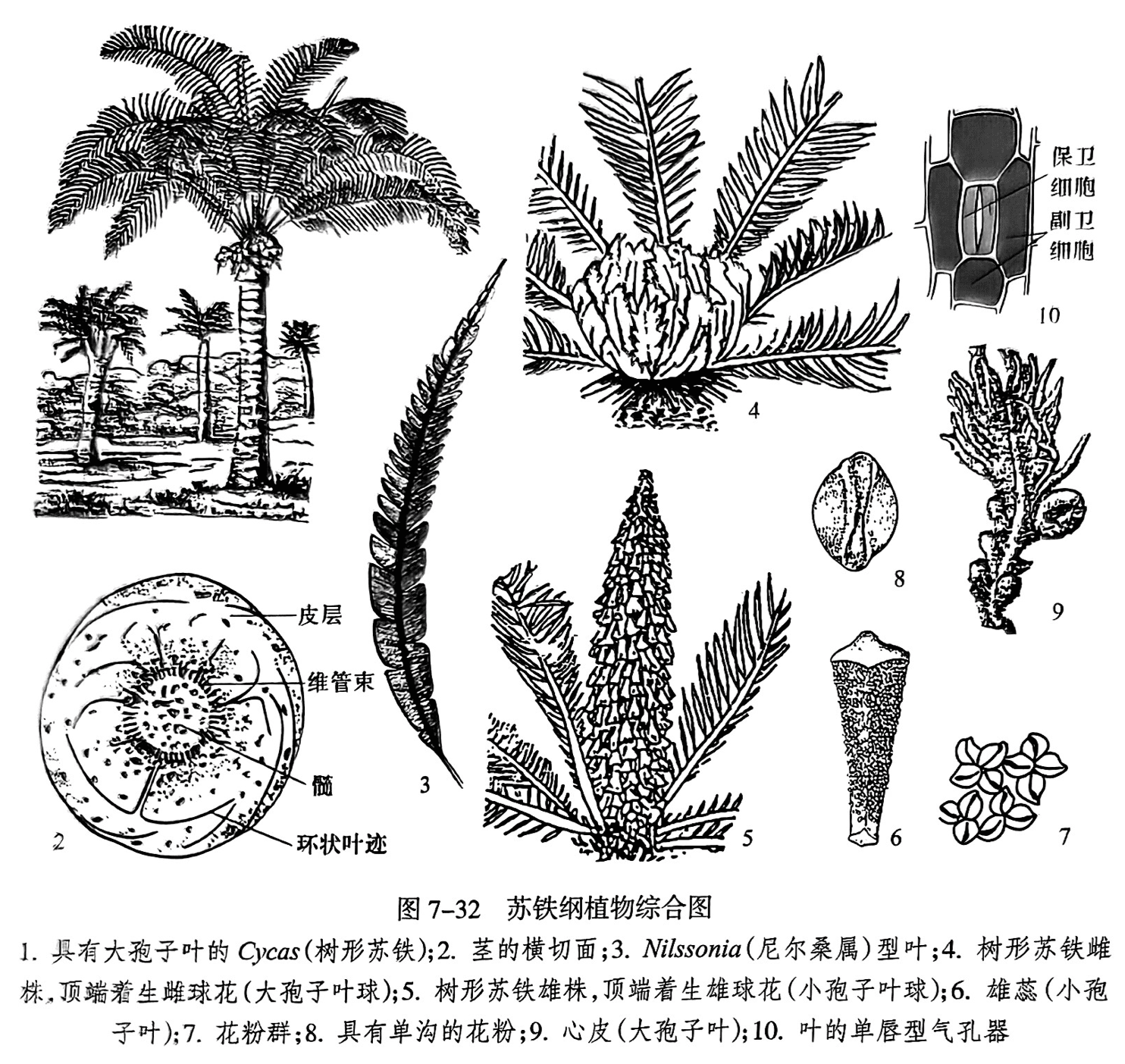
(1)苏铁纲(Cycadopsida)。本纲雌雄异株,球花单性;叶的形态与蕨类植物相似,为大型羽状复叶,幼叶亦呈拳曲状;大多数为平行脉或放射脉。叶坚硬,革质,表面细胞壁直,气孔器为单唇式(hoplocheilic),也称苏铁式(图7-32之10)。分布时代为石炭纪至现代,代表性化石如Nilssonia(尼尔桑)(图7-32之3)。
(2)本内苏铁纲(Bennettiopsida)。本纲又称拟苏铁纲(Cycadeoidopsida),雌雄同株,孢子叶球或两性或单性,有一些具有与被子植物相近的两性“花”;表皮细胞壁弯曲,气孔器为连唇式(图7-33之1d),分布时代为二叠纪至白垩纪。根据茎的分枝程度及生殖器官的构造,可将其分为拟苏铁科(Cycadeoidaceae)和威廉姆孙科(Williamsoniaceae)。
拟苏铁科的茎粗短,呈椭球形或圆柱状,高度不超过1m,多数不分枝,茎上覆永久性叶基及小鳞片。两性“花”长在叶腋间,在一条较粗的轴上,最外面为密螺旋状排列的羽状营养苞片,苞片上密覆细毛,苞片之上环状排列着基部联合的雄蕊(小孢子叶),轴的顶端为“雌花”(大孢子叶球)。叶化石为 Zamites(似查米亚)(图7-33之1c),呈螺旋状分布在树干的顶端。
威廉姆孙科的茎纤细,有的类型有二歧式分枝的枝(图7-33之2a),高可达2m,树干表面具有永久性排列的叶基。大、小孢子叶球常具有柄,着生于叶腋或枝顶,或为单性花或为两性“花”,成熟时开放。叶部化石常见3个属:①Ptilophyllum(毛羽叶),裂片线形,基部全部着生于羽轴腹面,斜伸,脉平行或呈放射状(图7-33之2c);②Pterophyllum(侧羽叶),叶呈单羽状,裂片基部全部附着于羽轴的两侧,平行脉;③Otozamites(耳羽叶),裂片基部收缩成耳状,放射脉。
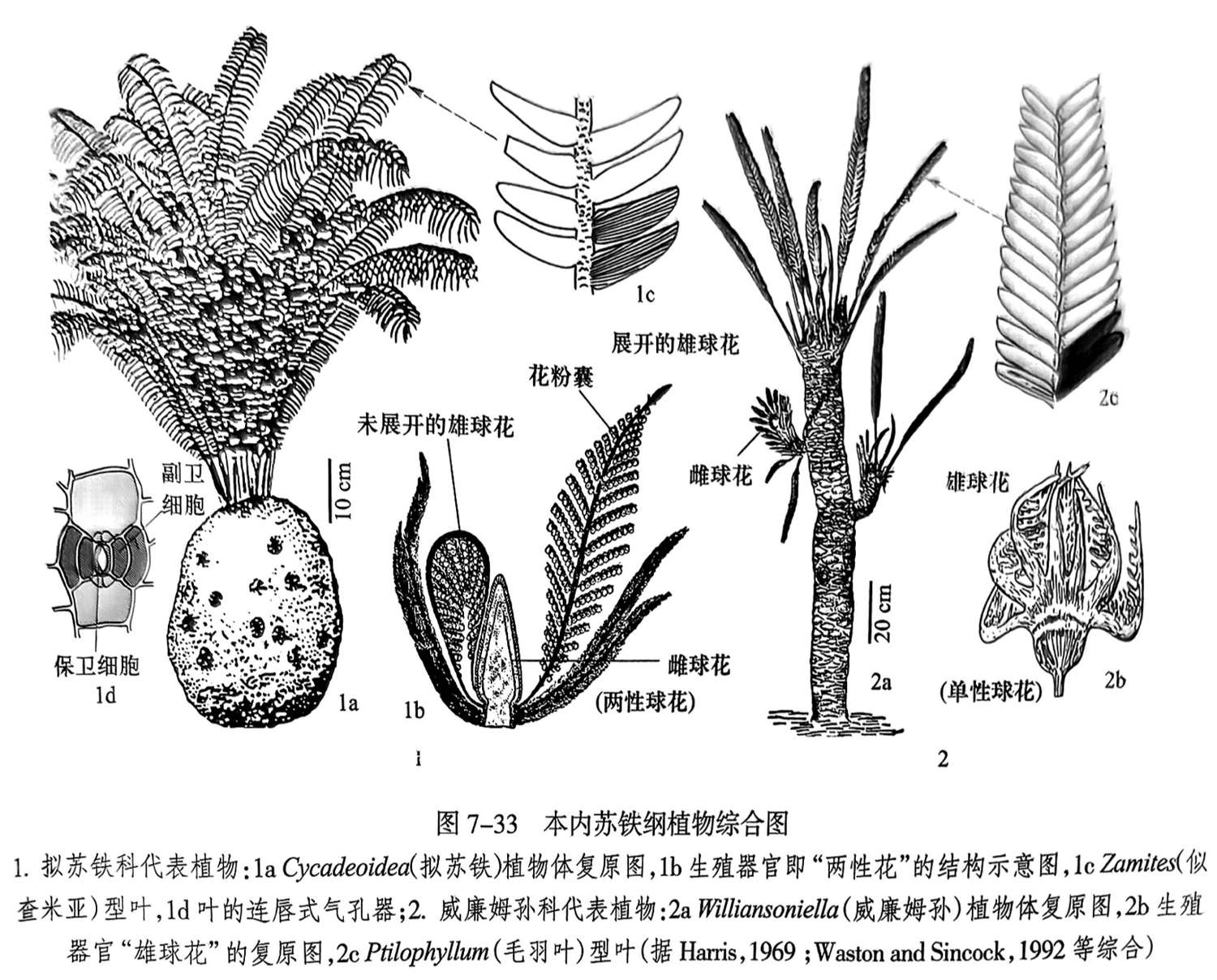
3.地史分布及生活型
苏铁植物门在晚古生代就已存在,在晚三叠世在各古大陆已广泛分布,在侏罗纪和早白垩世繁盛,遍及全球,甚至出现在南、北极。现代,本门植物退居很次要地位,有11属近219种,仅分布于热带及亚热带地区。现代苏铁植物的植株大多茎干粗壮、直立不分枝,但在早白垩世以前苏铁植物是以细茎类型为主的,这可能与其生活环境相关。我国华北发现于下二叠统细茎类苏铁,其共生植物及其围岩分析结果表明,它们是生活于热带暖湿的泛滥盆地或地势稍高的天然堤环境中的灌木型植物。苏铁植物在中生代极盛时期,很可能生活型已发生分化。至少自早白垩世开始苏铁植物已逐渐改变了以细茎类型为主的形式。这种变化可能与中生代晚期大范围的古气候变迁相关,使苏铁植物在退行性进化过程中存在分枝逐渐缩减而发展为单柱不分枝的趋向,形成在热带、亚热带环境中能适应干季的粗壮、不分枝的柱状植株。
四、银杏植物门(Ginkgophyta)
1.一般特征
现代银杏(Ginkgo biloba)为高达30余米的乔木,直径最大可达2m以上;雌雄异株;单轴式分枝,有长、短枝之分。在长枝上螺旋状着生单叶,在短枝上叶3~6枝呈密集螺旋状簇生(图7-34之1b、1c)。茎为内始式真中柱,次生木质部发达而髓不发育,年轮明显。叶具有长柄,呈单叶、扇形、肾形或宽楔形,或分裂成细长的裂片(图7-34之3~6)。自叶的基部伸出两条脉,然后多次二歧式分枝形成扇状脉。在同一植株上叶形有变化,在长枝上的银杏叶仅在叶中央有一浅裂口。而在短枝上及树干基部的根出叶,常多次深裂,经过化石的验证,认为这是一种返祖现象,也可称为进化异叶性,说明银杏植物在进化过程中,叶由分裂次数多、深裂向分裂次数少、浅裂的方向演化(图7-34之1e)。

2.分类
现代银杏为单种属的孑遗植物,本门分类主要依据化石资料。在地史时期银杏植物是一个类型多样而繁盛的“家族”。目前发现最早的银杏化石为2.7亿年前早二叠世的 Trichopity(毛状叶)。地史时期的银杏生殖枝和营养枝与现代银杏都存在比较大的差别。现代银杏在成熟时生殖器官上一般只有一枚无珠柄的种子,叶片为全缘,具有长柄;而地史时期的一些种类的银杏的生殖器官为有珠柄的多枚种子,叶片分裂深。化石记录表明银杏植物大致经历了雌性生殖枝减退。珠柄最终消失,胚珠减少并增大,营养叶(或苞片)趋向蹼化,叶柄从无到有的演化过程。由于雌性生殖枝化石资料相对比较少,本门分类主要依据营养枝叶片化石资料,分为两个目:银杏目和茨康目。
(1)银杏目(Ginkgoales)。叶具有柄,呈扇形、肾形或楔形。为扇状脉。叶常二歧式分裂成2-8个或更多个的最后裂片。单叶脱落,有两条叶脉自叶基部进入叶片,在叶基的断面上留有双叶迹。化石代表如 Ginkgoites(似银杏)、Baiera(拜拉)、Sphenobaira(楔拜拉)等(图7-34之2~4)。
(2)茨康目(Czekanowskiales)。叶窄长,多数无柄,簇生于短枝上,简单或二歧式分枝,每束叶(5~18个)与鳞片包着的短枝一起脱落,基部有一条脉进入叶片后分叉(图7-34之6)。目前对于这类叶的自然分类归属还存在很多疑问,本教材依照传统观点将其归入银杏类。
3.地史分布及生态
银杏植物始见于晚古生代,较可靠的化石记录为二叠纪,在中生代银杏类型多样,尤其是在侏罗纪和早白垩世达到极盛阶段,广泛分布于亚欧大陆的温带植物地理区。在早白垩世晚期突然衰退,新近纪地理分布已缩小,但在中欧尚有银杏类分布。自第四纪冰期后,银杏植物在中欧、北美等地全部灭绝。现存的银杏仅一属一种,广泛栽培于世界各地,但野生状态的银杏目前仅发现于我国浙江西部天目山。银杏植物主要分布于温带区,为中生生态类型。
五、松柏植物门(Coniferophyta)
1.一般特征
松柏植物是现代最繁盛的裸子植物,为高度分枝的乔木或灌木,茎为真式中柱,无叶隙,次生木质部中薄壁细胞少,致密,初生木质部内始式,管胞径向壁上具有缘纹孔。叶多为针状、锥状和条状,呈螺旋形排列。雌雄同株或异株,球果单性。雄性球果(小孢子叶球)的轴上螺旋状着生雄蕊(小孢子叶),每个雄蕊上有花粉囊2~15个;雌性球果绝大多数由具有种子的鳞片绕轴紧密排列而成。
2.分类
本门一般分为科达纲和松柏纲。
(1)科达纲(Cordaitopsida)。本纲已灭绝。植物体为茎干直径不超过1m的细高乔木,茎干基部有的具有高位支持根(图7-35之1)。树干上部分枝较多,组成较大的树冠。单叶,螺旋状着生于枝,叶无柄,为带形或舌形(图7-35之2),大小不一,长者可达1m,短者仅几厘米,平行脉,平行脉之间有间细脉(interstitials)。茎干皮部厚,髓部大,由于髓部薄壁细胞分裂不连续,保存为髓模化石时其表面具有横列粗纹。常见化石为 Cordaites(科达)。
本纲始见于晚泥盆世,在晚石炭世至中二叠世最繁盛,遍及全球各植物地理区,在热带与鳞木等同为重要的造煤植物,在三叠纪有少量残存,在三叠纪后期灭绝。
(2)松柏纲(Coniferopsida)。本纲除少数为灌木外,绝大多数为常绿乔木,为单轴式分枝,树冠高大,如产于北美的巨杉(Sequoiadendron)高达91m,直径达8~10m,这种巨杉由于寿命极长,被人们称为世界爷。叶为小型,常呈鳞片形、锥形、针形、条形或披针形等,大多数为单脉,因而又称之为针叶树。叶在枝上的排列方式多样,有簇生、螺旋状着生、轮生、交互对生、假两列着生等(图7-36),化石中还发现叶呈鞘状围于枝上(图7-36之8)。叶片角质层厚,气孔下陷。生殖器官绝大多数为单性的球花,雌雄同株,极少数异株。本纲进一步分为:伏脂杉目(Voltziales)、松柏目(Coniferales)、紫杉目(Taxales)和苏铁杉目(Podozamitales)。
伏脂杉目:具有古生代科达纲向现代松柏纲过渡的特点。雌、雄球花都由若干生殖短枝绕轴着生于苞片腋部组成,雌、雄生殖短枝呈切向平展,其上着生的鳞片中有的部分具有胚珠,胚珠数目不一,胚珠形态呈直立或倒转状。大多数具有螺旋状着生的鳞片状叶,其中掌鳞杉科是中生代有别于松柏植物门其他科的重要类群(图7-37之1),以产花粉 Classopollis(环沟粉)而闻名。见于晚石炭世至白垩纪。
松柏目:是现生最多的松柏纲类群。除罗汉松科为单个种子外,其他各科的雌性生殖器官都为球果。其中 Metasequoia(水杉)是典型代表,叶呈假两列状,球果的果穗为交互对生(图7-37之2),它是在先有化石植物研究多年后,才在我国湖北利川等地发现了现生孑遗代表,故称为“活化石“,本目始现于三叠纪,在侏罗纪变得多样化,在现代广布全球

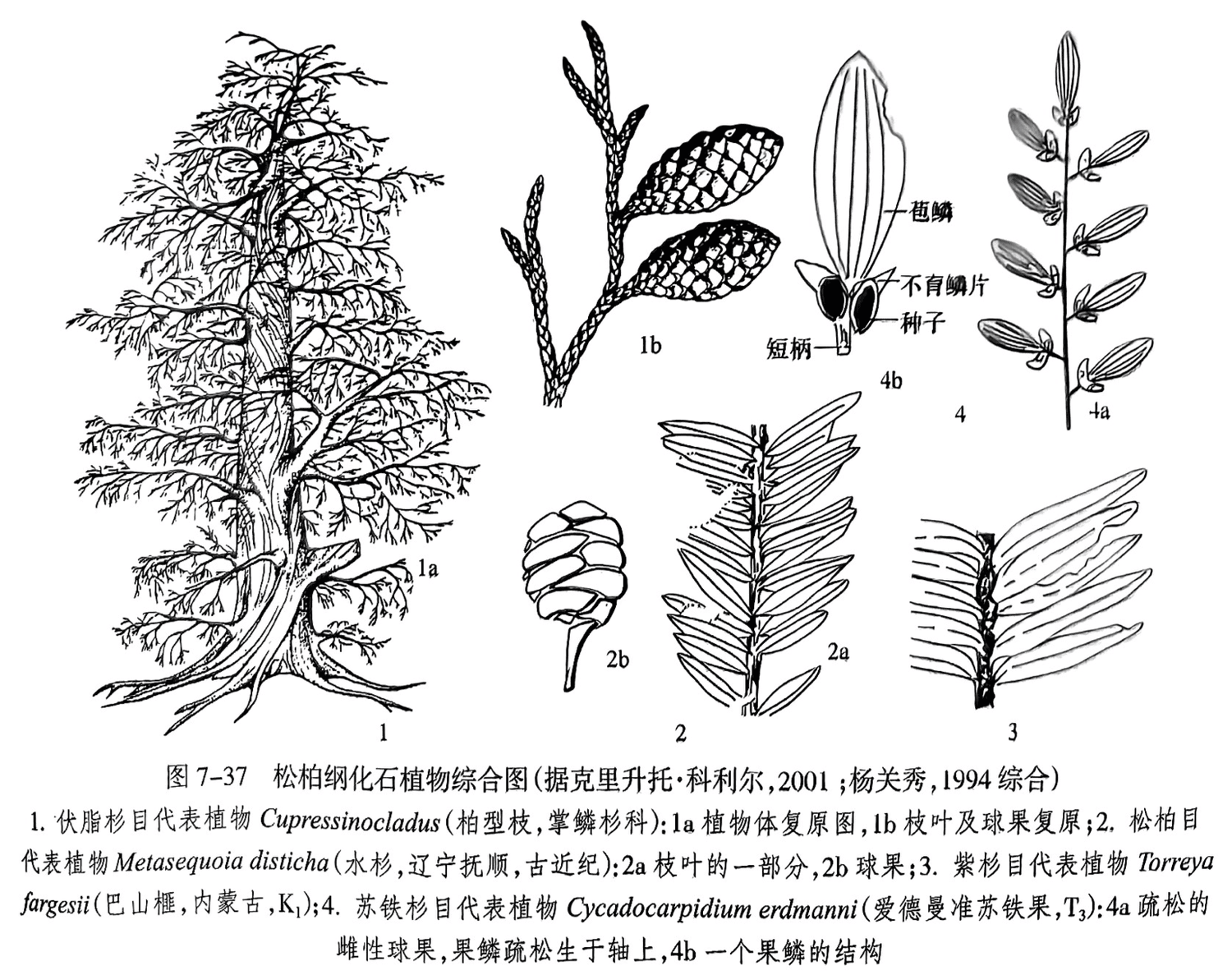
紫杉目:为常绿灌木或小乔木、具有鳞芽(scalybud),为螺纹管胞。叶为披针形或条形,基部变窄或具有小叶柄,螺旋生长,因叶柄扭转而成两列,中脉凹陷,为单面气孔型,叶背面中脉两侧各具有一条纵向排列的气孔带,雄果穗单生。代表化石如 Torreya fargesil(巴山榧)(图7-37之3)、Cephalotaxopsis(拟粗榧)等。本目始见于三叠纪,在白垩纪化石最为多见,现代仅分布于北半球。
苏铁杉目:枝叶化石以 Podozamites(苏铁杉)最为常见,枝轴细,叶稀,呈螺旋状着生,具有弧形脉。相关的生殖器官化石为 Cycadocarpidium(准苏铁果)(图7-37之4),叶状的苞片下有两枚种子,是晚三叠世的标准化石.地质时代为三叠纪至早白垩世。
3.地史分布与生态
本纲始见于晚石炭世早期,在中生代全面繁盛,在新生代松柏纲是裸子植物中生存最多的类群。现代松柏纲植物广布于不同纬度和不同海拔高度的平原、山区,常形成大片的针叶林。
六、买麻藤植物门(Gnetophyta)
1.一般特征
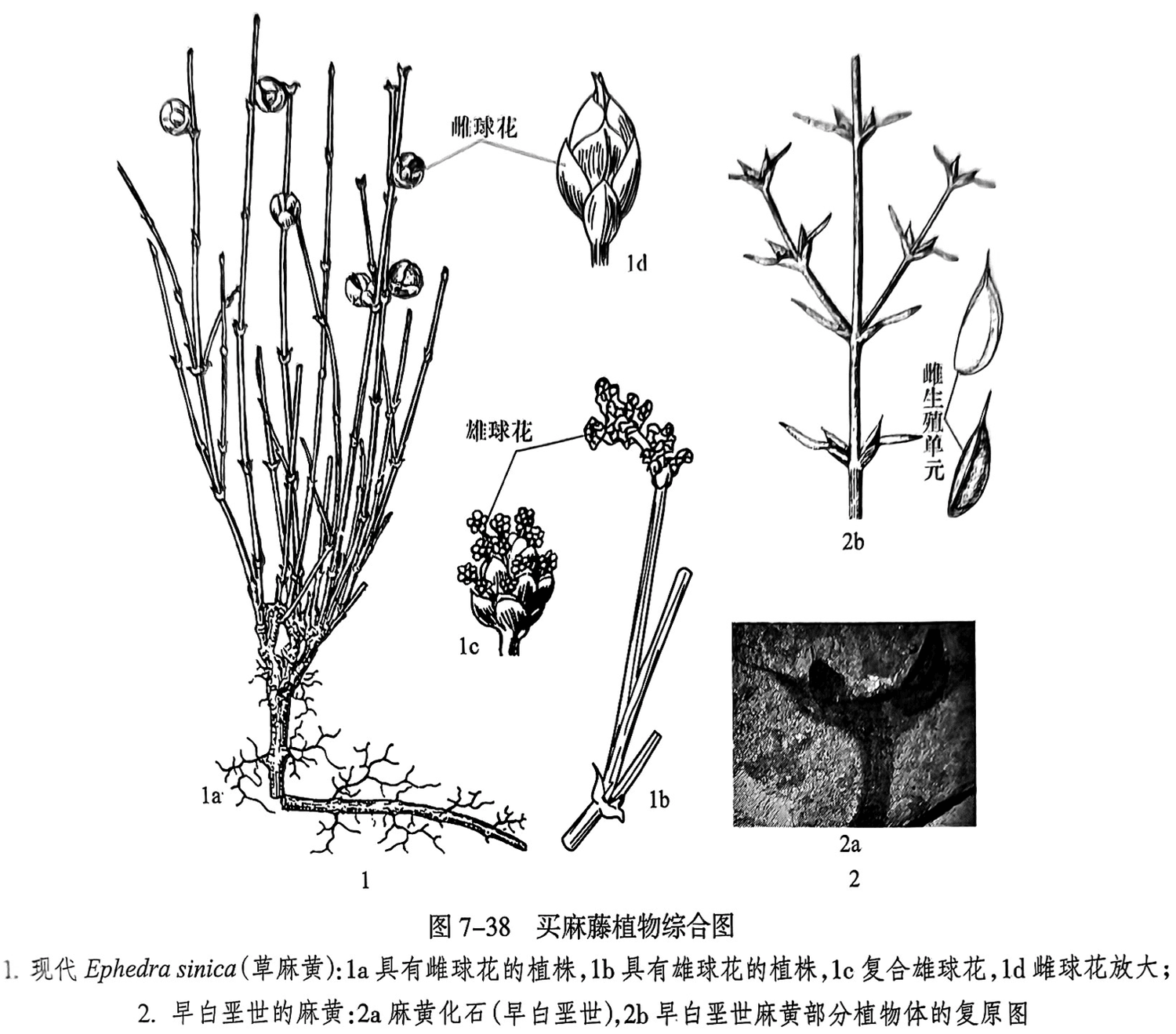
买麻藤植物是植物分类体系中比较特殊的一类,它与其他裸子植物和被子植物有比较大的差异。买麻藤植物有灌木、木质藤本和草本状小灌木类型。次生木质部常具有导管,无树脂道。叶为对生或轮生,叶片种类多样,有的为细小膜质,呈鞘状或绿色扁平状(似双子叶植物叶),也有的为肉质,呈带状(似单子叶植物叶)。球花为单性,或异株或同株,球花(孢子叶球)有类似于花被的盖被,也称假花被。盖被膜质、革质或肉质;胚珠有1枚,珠被有1~2层,具有珠孔管(micropylar tube);精子无鞭毛,颈卵器或极其退化或无;成熟雌球花为球果状、浆果状或细长穗状(图7-38之1)。
2.分类
买麻藤植物由于茎具有导管,无颈卵器,苞片特化成花瓣状等,显示了被子植物的某些特征,因此它们被认为是裸子植物和被子植物之间的“过渡类型”,甚至有的学者认为买麻藤植物是被子植物的祖先。买麻藤植物门包括三大独立的类群,但这三大类群之间并无密切的亲缘关系,现生植物学将它们归为3个独立的科:麻黄科、买麻藤科和百岁兰科。
3.地史分布与生态
买麻藤植物门的麻黄类植物在早白垩世地层中就有较多产出。在中国东北早白垩世地层中发现了大量的麻黄类化石,如古斑子麻黄、小前麻黄(图7-38之2)、洪涛麻黄等。现代买麻藤植物麻黄和百岁兰主要生活在比较干旱的地区。而在早白垩世地层中发现的买麻藤植物生活于热带和亚热带森林。