4 三叶虫
内容节选自《古生物学》(童金南主编,2021年版)第五章 古无脊椎动物 第四节 三叶虫动物
一、 一般特征
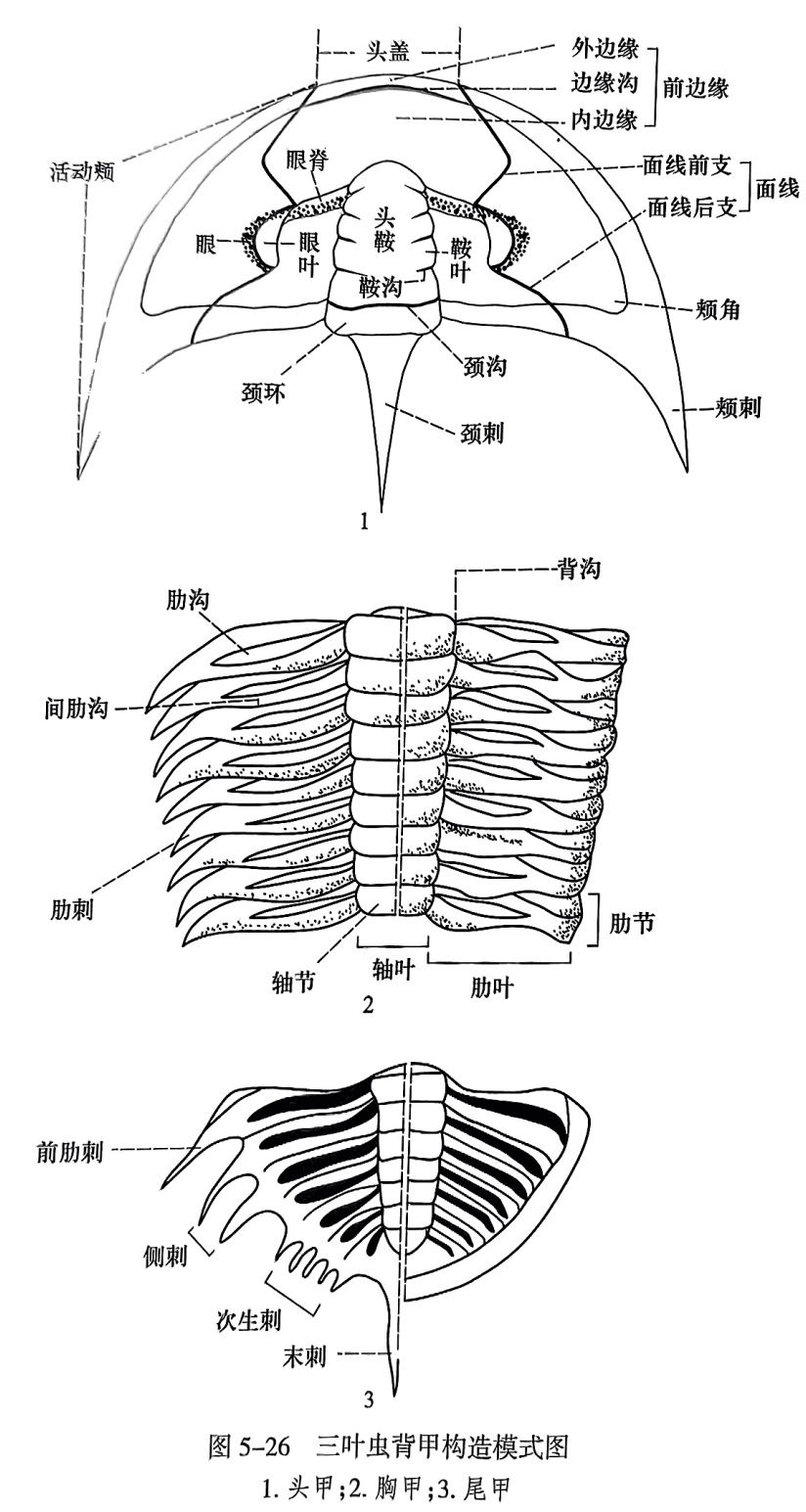
三叶虫(trilobites)是节肢动物门中已灭绝的一个纲,属于节肢动物门(Arthropoda)三叶形亚门(Trilobitomorpha)三叶虫纲(Trilobita),化石仅见于古生代海相地层中。三叶虫身体扁平,奔侧披以坚固的甲壳、腹侧为柔软的腹膜和附肢。背甲(dorsal shield)在纵向上被两条背沟(domsalfarmow)分成三部分:中间部分为轴叶(axial lobe),两侧部分为肋叶(pleural lobe),因而得名三叶虫。三叶虫自前而后又横向分为头部(头甲)、胸部(胸甲)和尾部(尾甲)(图5-26)。
二、三叶虫的背甲构造
三叶虫背甲成分以碳酸钙和磷酸钙为主,质地坚硬,是地史时期最早大量保存的化石。背甲主要包裹着虫体的背侧和腹侧边缘,呈长卵形或椭圆形,略上隆;通常长3~10cm。小者不及6mm,大者可达70cm。
(一)头甲
头甲(cephalic shield 或 cephalon)构造复杂,是分类和属种划分的主要依据。多呈半椭圆形,中间有隆起的头鞍(glabella,复数为 glabellae)和颈环(occipital ring),其余称颊部(cheek),颊部中央常具有眼和眼叶(palpebrallobe),多数三叶虫通过眼的内侧与眼叶之间有一对狭缝切穿颊面,将头部分为头盖(cranidium, cranidia)和活动颊(free cheek, librigena),这对狭缝称面线(facial suture)(图5-26之1)。
(1)头鞍。头鞍是头部中间隆起部分,两侧为背沟所限,其形状多样,有两侧平行的,有向前收缩的,有中部收缩的,也有向前扩大的,等等。其向上隆起的程度也各异,一般适度上凸,有的凸度极大呈球形,有的则低平而难以与颊部区分。头鞍有时光滑,有时具有成对的沟,为头部分节的痕迹,称头鞍沟或鞍沟(glabellar furrow)。鞍沟数最多不超过5对,可在中部连接,通常自前向后加长、加深。鞍沟把头鞍分成若干鞍叶(glabellar lobes)。在某些类群中头鞍中后部中央有一小瘤称为中瘤(median node)。头鞍之后为颈环,其间以颈沟(occipital furrow)为界。颈环中部可具有颈瘤(occipital node),有时向后延伸出颈刺(occipital spine)。
(2) 前边缘(front area)。前边缘是头鞍之前背壳的总称,被边缘沟(marginal furrow)区分为外边缘(anterior border)和内边缘(brim)。其发育程度与头鞍向前延伸的程度成相互消长的关系。
(3)固定颊(fixed cheek或 fixigena)。三叶虫的颊部如果具有面线,面线之间包括头鞍在内的区域则统称为头盖,两侧称为活动颊。在面线内侧头盖外缘有一对半圆形的隆起部分,称为眼叶,对眼起支持作用。其形状、长短、宽窄,以及和头鞍的相对位置十分重要。眼叶前端可具有一条上凸的脊线与头鞍前侧角相连,称为眼脊(ocularridge)。在眼脊之后,眼叶与头鞍之间的范围称为固定颊眼区(oculararea)或固定颊(狭义)。在眼脊之前,头鞍两侧的背沟向前延伸线与面线前支之间的范围称为眼前翼(preocular limb)。在固定颊眼区之后,面线后支与背沟之间的范围称为后侧翼(posterolateral limb)。人们又经常把眼前翼、固定颊眼区(狭义的固定颊)和后侧翼合称为固定颊(广义)。实际上我们在描述固定颊宽度时,仅指固定颊眼区的宽窄。
(4)活动颊及眼。活动颊(或自由颊)是指面线外部的颊面,眼位于其内缘,并可分为两类,即复眼和聚合眼:前者为一个眼角膜覆盖所有的小眼体,为多数三叶虫所具有;后者每个小眼体各自为一个眼角膜覆盖,小眼体之间为巩膜板所隔,仅为镜眼虫类和少数手尾虫类具有。头甲侧缘与后缘之间的夹角称为颊角(genal angle),它可向后伸长成颊刺(genal spine)。
(5)面线类型。面线是通过眼与眼叶之间切穿颊部的狭缝。我们将眼叶之前的面线称为面线前支,其后的面线称为面线后支。面线前支可在头鞍之前的背侧或腹侧会合,亦可在前缘会合。面线后支根据伸展方向分为后颊类面线(opisthoparian suture)(后支在颊角后方)、前颊类面线(proparian suture)(后支在颊角之前)和角颊类面线(gonatoparian suture)(后支切于颊角)。有的处在头甲边缘,称为边缘类面线(marginal suture)(图5-27)。也有的三叶虫无面线。
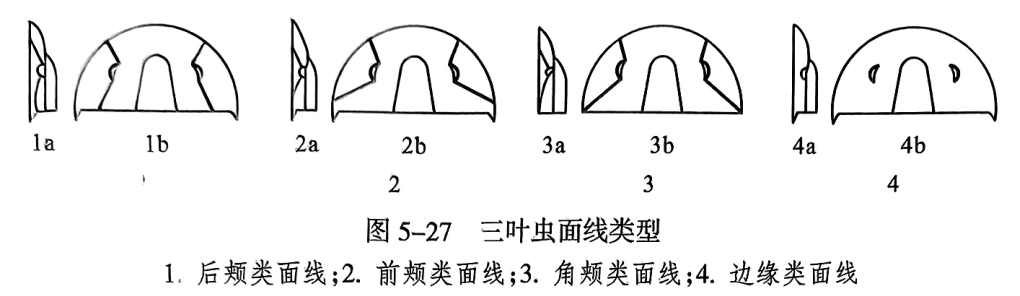
(二)胸甲
胸甲(thoracic shield或 thorax)由若干形状相似、互相衔接并可活动的胸节组成。关节处可弯曲,因此身体能卷曲。胸甲中间隆起部分为轴叶,两侧为肋叶(图5-26之2)。轴叶上轴节之间以半环(half ring)和关节沟(articulating furrow)相互衔接,只有身体卷曲时才能见到。肋部各肋节之间为间肋沟(interpleural groove)所隔;肋节上的沟称为肋沟(pleural furrow);肋节末端钝圆,或延长成肋刺(pleural spine);有的在肋节末端的前方有一个斜面,称为关节面(articulating facet)。
(三)尾甲
尾甲(pygidial shield或 pygidium)多呈半圆形,由若干体节融合而成,其节数自1至30不等。仅第一个轴节具有半环和关节沟,以此和胸甲衔接。肋沟一般较间肋沟深而宽,有时两者都不发育。尾甲边缘的宽度因属而异,有的还具有尾刺。根据刺和肋节的关系,以及所处位置,又可将其分为前肋刺、侧刺、末刺和次生刺(图5-26之3)。
尾甲和头甲大小比例关系是三叶虫分类的另一条重要依据,据此,可将尾甲分为4种类型:①尾甲极小的称小尾型(micropygous);②尾甲略小于头甲的称异尾型(heteropygous);③尾甲与头甲大致相等的称等尾型(isopygous);④尾甲大于头甲的称大尾型(macropygous)。
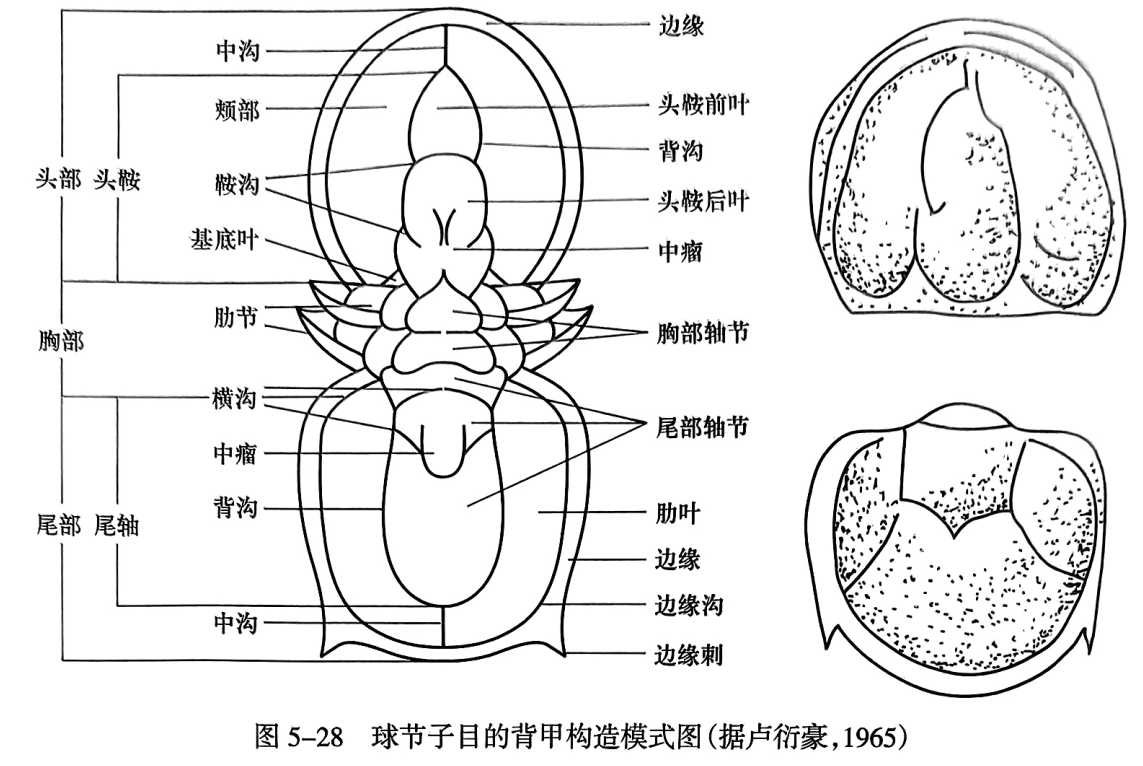
(四)球接子目的背甲构造
球接子目(Agnostina)的头部无眼及面线,个体比较小,其背甲基本构造与前述三叶虫的背甲构造有一定的区别,具体构造名称见图5-28。
三、三叶虫的分类
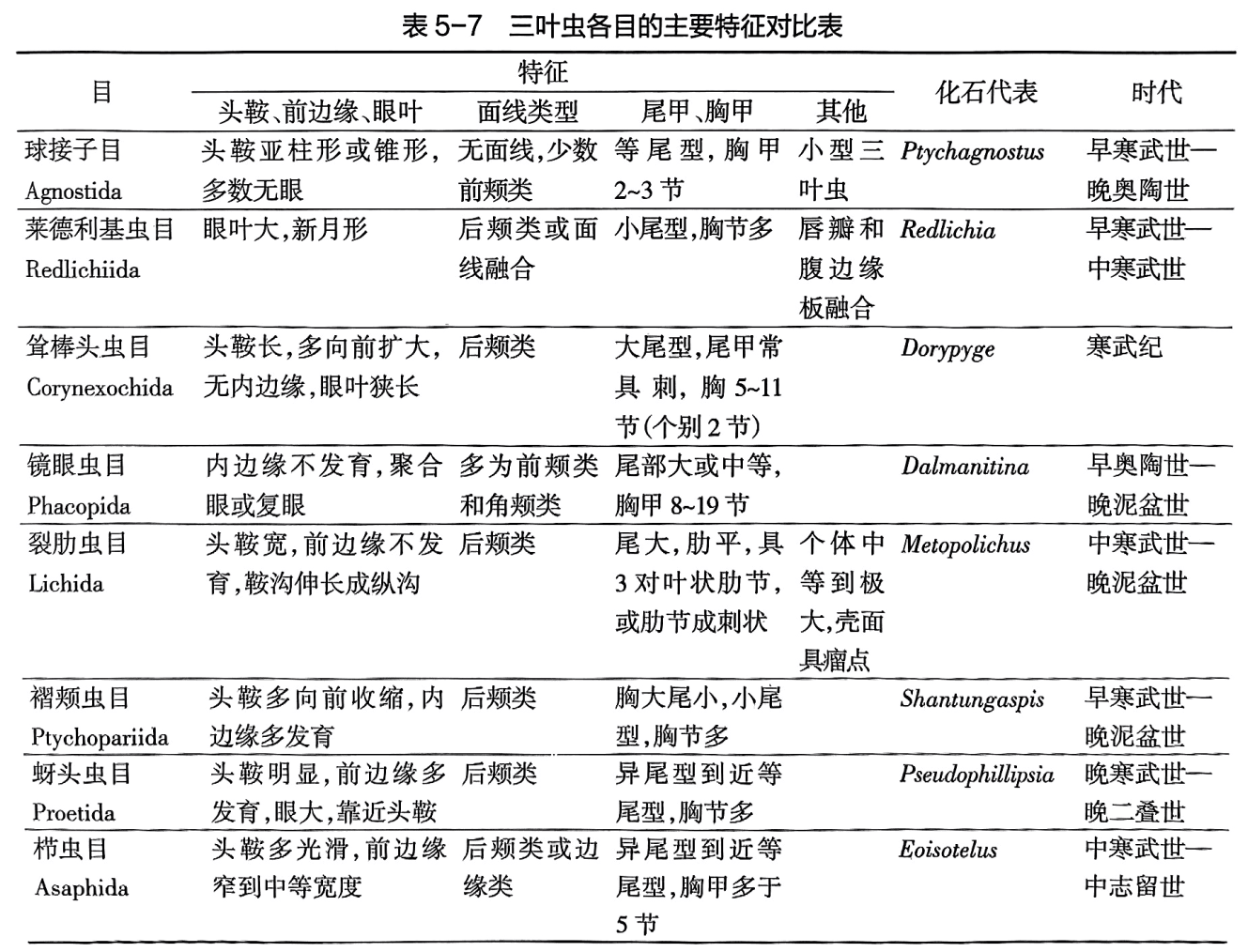
由于三叶虫早已灭绝,其内部构造和腹部、附肢的化石资料又非常稀少,属种个体发育的研究也不多,以及其他一些原因,所以各家对它们的分类方案分歧甚大。目前趋向于利用综合特点进行分类,即考虑三叶虫的主要形态特征,如头甲轴部构造、面线类型、胸甲和尾甲特点,以及个体发育、唇瓣特征等。1997年修订版的《无脊椎动物古生物学论著》(Treatise on InvertebratePaleontology)将三叶虫纲分为8个目,其中主要是将原归入褶颊虫目的部分类别分解出来。各目的主要特征列于表5-7。
四、三叶虫的生态与形态功能分析
三叶虫在二叠纪末完全灭绝,因此要了解各类三叶虫的生活方式和解释其各种构造的功能。只有对三叶虫的身体结构构造的形态功能、它们生活时的无机和有机环境,以及它们的分布和保存状况进行综合分析。才能得到比较正确的认识。
三叶虫化石常与海生动物如珊瑚、海百合、腕足动物、头足动物等化石共生,同时三叶虫化石只在海相沉积岩中出现,在陆相地层中从未被发现过,因而证明它是海生的。它们大多适于浅海底栖爬行或半游泳生活;另一些种类可在远洋中游泳或漂游,以原生动物、海绵动物、腔肠动物、腕足动物的尸体,或海藻及其他细小生物为食。
多数三叶虫的身体扁平,背部披以硬甲,腹部为柔软的腹膜和可爬行的步足(即分节、末端具爪的内肢)。眼位于头甲背侧,这就是腹部贴伏海底生活的特点;扁平的身体和大量的刺可增加浮力;加上鳃状外肢有助于游泳,因此三叶虫还具有一定的游泳能力。可是三叶虫缺乏流线型的体形,又无良好的游泳器官,所以它只能是一种不高明的游泳者,如褶颊虫目、莱德利基虫目等。
由于辐射适应,三叶虫的某些类群可钻入泥沙中生活。其头部结构极为坚硬,无眼,曾在装饰昂尾虫(Onniaornata)的消化系统中发现大量泥质物,证明这类三叶虫是从泥土中摄取养料的,如同现代的蚯蚓。有的三叶虫头甲愈合,复眼靠近头鞍或无眼,尾小,末端尖锐成刺状,如Olenellaus(图5-29之1),它与现代海洋中的鲎 Limulus 十分相似。后者在沿岸沙质海底上用锐利的头部前缘和附肢挖掘淤泥,用刺状尾插入泥沙,使身体慢慢向前移动。另外,适于在松软的泥沙或淤泥海底生活的三叶虫,如多角虫(Ceratarges)(图5-29之3)的肋刺和尾刺都很发育,使之不至陷入淤泥中,而且多角虫具有柄状眼,这是防止淤泥影响视线,并扩大视域的进化结果。

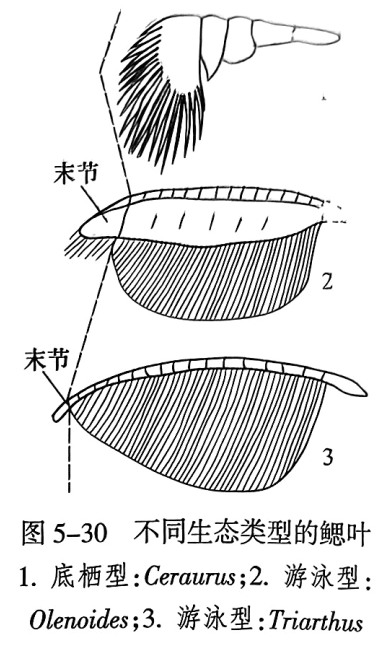
还有个体小,头鞍膨大,眼极其发育,位于头部边缘,甚至延伸至腹侧的三叶虫,如奥陶纪的环尾虫(Cyclopyges)(图5-29之4)和黏合盾甲虫(Symphysops)(图5-29之5),代表远洋区游泳的类型。个体小,带长刺的三叶虫,也多营漂浮生活,如刺壳虫(Acidaspis)(图5-29之6)。外肢上鳃叶又长又大如刷子和船桨的三叶虫,如 Olenoides(图5-30之2)和 Triarthus(图5-30之3)等,其分布多为世界性的,这说明它们善于游泳或漂游;而外肢上鳃叶不发育的三叶虫如 Ceraurus(图5-30之1),其分布仅限于局部地区,它们属于底栖生活类型。
球接子是一种特殊的三叶虫,营远洋漂游生活。这是因为:①其壳体小,结构简单,腹边缘极窄,唇瓣呈强烈拱凸的格架状,因此体轻,利于漂浮;②无眼是由于被动漂浮的结果;③呈世界性分布;④大量出现于含黄铁矿的黑色页岩或灰岩中,这种岩性很少含有正常的底栖生物化石,反映的是海水较深,水流滞缓,正常底栖生物不易生存的沉积环境,这些都是远洋生物的特点。有人还认为球接子生活时多处于卷曲状态,头和尾互相拍打着活动。
三叶虫的幼虫凸度较大,往往具备较发育的刺,说明幼虫阶段营漂浮生活。
此外,从奥陶纪开始出现的大量的肉食动物(鹦鹉螺、原始鱼形动物等);它们直接威胁三叶虫的生存。为了防御敌人的侵害,许多三叶虫如 Nileus(宝石虫)、Cyclopyge(圆尾虫)等的尾甲增大,提高了游泳速度。同时头尾嵌合使整个身体卷曲成球形、盘状,以保护柔软的腹部,并可迅速跌落或潜伏至海底,以逃避敌人的进攻。
五、三叶虫的演化与地史分布
在寒武纪早期三叶虫就已出现,在8个目中已出现5个目,属种数量也很丰富,遍布世界各地,并已显示出生物分区的特性,因此古生物学家都认为三叶虫的远祖早在寒武纪之前就已存在。从三叶虫化石记录可以看出三叶虫形态与构造在地质历史上的四点演化趋向:
(1)在早寒武世三叶虫总的形态与构造特点是头大、尾小、胸节多;头鞍长、锥形、鞍沟显著;眼叶发育,靠近头鞍;胸节肋刺发育。目前我国已发现的早寒武世三叶虫有200余属,以菜德利基虫目最重要,如 Redlichia、Palaeolenus等;在早寒武世晚期褶颊虫目中的某些类群开始出现,如小遇仙寺虫(Yuehsienszella);这个时期也有耸棒头虫目的代表,如湖南头虫(Hunanocephalus)、似手尾虫(Cheiruroides)等,但数量还比较少。
(2)在中、晚寒武世三叶虫发生辐射适应,这个时期三叶虫总的形态与构造特点演变为尾甲加大,多为异尾型;胸节数减少,头鞍较短,多具内边缘,眼叶较小;鞍沟数量减少,且很少穿越头鞍。属种数量变得更多,我国已描述的三叶虫属达500余个。在寒武纪中期莱德利基虫目只剩下个别代表。在寒武纪中、晚期褶颊虫目和耸棒头虫目居统治地位,如 Kaotaia、Anomocarella、Damesella、Blackwelderia、Dorypyge、Chuangia等;此外球接子目在许多地区大量地出现,如Ptychagnostus、Pseudagnostus等;栉虫目有许多代表,如晚寒武世的 Yueshangia、Proceratopyge等;蚜头虫目也有少量出现,如晚寒武世的 Plethopeltis等。
(3)在奥陶纪时、三叶虫发生显著变化,在形态与构造上比寒武纪三叶虫的尾甲要大,多为等尾型甚至大尾型:胸节数量进一步减少,一般为8~9节;头鞍向前扩大,鞍沟、背沟,甚至颈沟都不发育。寒武纪的许多重要类别相继灭绝或衰退;而栉虫目有重要发展,如Eoisotelus、Asaphellus等。耸棒头虫目也非常丰富,如 Illaenus等。但奥陶纪早期的三叶虫特点与寒武纪后期颜相似。如 Asaphopsis、Dactylocephalus。这时由于呈世界性分布的笔石动物繁盛,活动力强的肉食动物头足纲兴起,三叶虫在海洋中不再是居统治地位的生物,地层划分除三叶虫外,笔石和头足纲也具有极重要的地位。特别在世界性地层对比中,后两者似乎更为重要。
(4)在志留纪至二叠纪,三叶虫急剧衰退,只留下少数类别,主要为镜眼虫目、褶颊虫目和蚜头虫目的少数代表,如 Coronocephalus(中志留世)、Dalmanitina(中奥陶世至早志留世)、Aristoharpes(优美镰虫)(志留纪)、Latiproetus(宽边砑头虫)(志留纪)、Phacops(镜眼虫)(志留纪和泥盆纪)、IFeberiphillipsia(韦氏菲氏虫)(早石炭世)、Pseudophillipsia(假菲氏虫)(二叠纪)等。它们的地层学意义不大。