十三、植物
(一)植物的观察要点
植物化石观察首先注意化石的保存特点。除少数茎干表面和内部髓腔形成的印模或核化石外、大多数是炭化作用形成的炭质压膜化石,其中多数是植物的枝叶化石,这时应重点观察叶的形态、结构、叶序、叶脉等特点,具体如下:
(1)对于石松植物门,注意区分叶座与叶痕。
(2)对于植物枝叶化石,注意观察小羽片基部形态和着生的位置。
(3)注意叶序、脉序的观察(如果叶脉保存完好)。
(二)代表性植物化石属描述
1.蕨类植物
(1)石松植物门
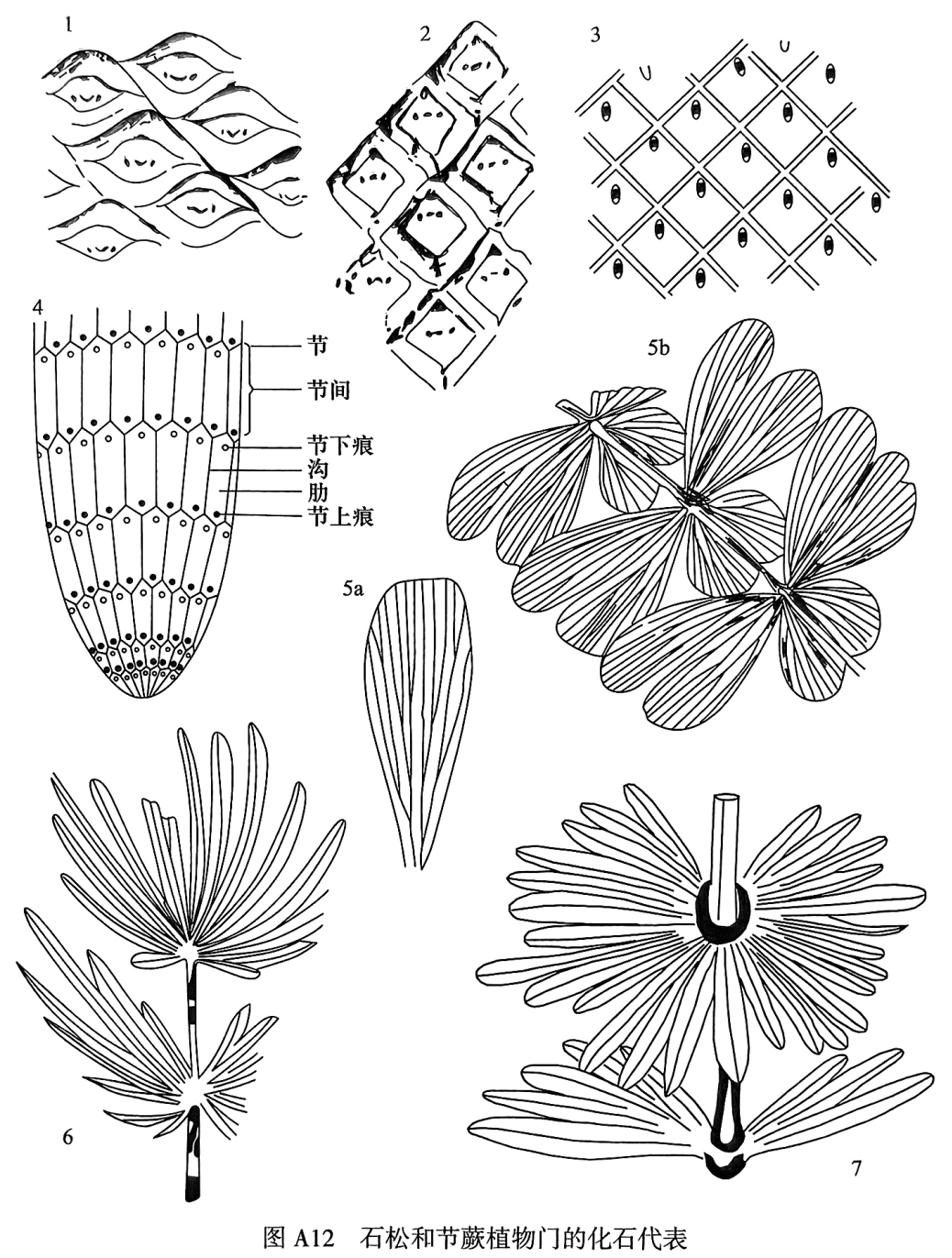
Lepidodendron Sternberg,1820(鳞木):是鳞木目的典型代表。叶座为菱形、长纺锤形等不同形态,在茎、枝上呈螺旋状排列;叶痕呈横菱形、斜方形等,中间具有束痕,两侧有通气道痕,紧靠叶痕顶端发育出很小的叶舌痕(图A12.1)。生活于石炭纪—二叠纪,分布于世界各地。
Cathaysiodendron Lee,1963(华夏木):叶座与叶痕形状相似,后者略小,为正菱形、菱形或横菱形,呈螺旋状排列;顶、底角略具有横皱纹。叶座间有凸起的、曲折的间隔带,使树干表面呈曲折状。束痕和侧痕位于叶痕侧角的连线上,叶舌穴位于叶痕顶角上端(图A12.2)。生活于晚石炭世一二叠纪,分布于华夏植物地理区。
Leptophloeum Dawson,1861(薄皮木):为乔木状,二歧式分枝;叶座大,呈菱形,表面光滑,为螺旋状排列;叶痕小,呈卵形,位于叶座中部或上部,叶痕中只有一条束痕,无叶舌;叶呈线形,单脉(图A12.3)。生活于中-晚泥盆世,分布于世界各地。
(2)节蕨植物门
Calamites Suckowi,1784(芦木):是一个广义的属名,既是代表古生代木贼目芦木类茎的髓模化石属名,又是古生代芦木植物体的总称。髓模表面纵肋和纵沟在节上交互排列(图A12.4),具有节下痕。在晚石炭世一二叠纪最盛,分布于世界各地。
Sphenophyllum Koening,1825(楔叶):为楔叶目的枝叶化石,叶轮生,数目常为3的倍数,多为6枚,每枚叶的基部收缩成楔形,叶脉为扇形(图A12.5a--b)。生活于晚泥盆世一二叠纪,在早一中二叠世最盛,分布于世界各地。
Lobatannularia Kawasaki,1927(瓣轮叶):是古生代东亚华夏植物地理区特有的芦木类枝叶化石。在末级枝上每轮叶有16~40枚;叶形状和着生方式同轮叶,但叶长短差别大,多少向外向上弯曲,呈明显的两瓣状;具有上、下叶缺,一般下叶缺明显,近叶缺处的叶最短。叶基部分离或大多数不同程度地连合。顶叶轮不分为两瓣,呈卵形或圆形(图A12.6)。生活于二叠纪,繁盛于晚二叠世。
Annularia Sternberg,1823(轮叶):是古生代木贼目芦木类的枝叶化石。枝对生,呈两侧对称。叶轮生,与末级枝几乎在一个平面上呈辐射状直伸排列。每轮叶有6~40枚,叶互相分离,大多数长短相等;叶形多种多样,呈线形、倒披针形或匙形等,单脉(图A12.7)。生活于晚石炭世一二叠纪,分布于世界各地。
(3)真蕨植物门
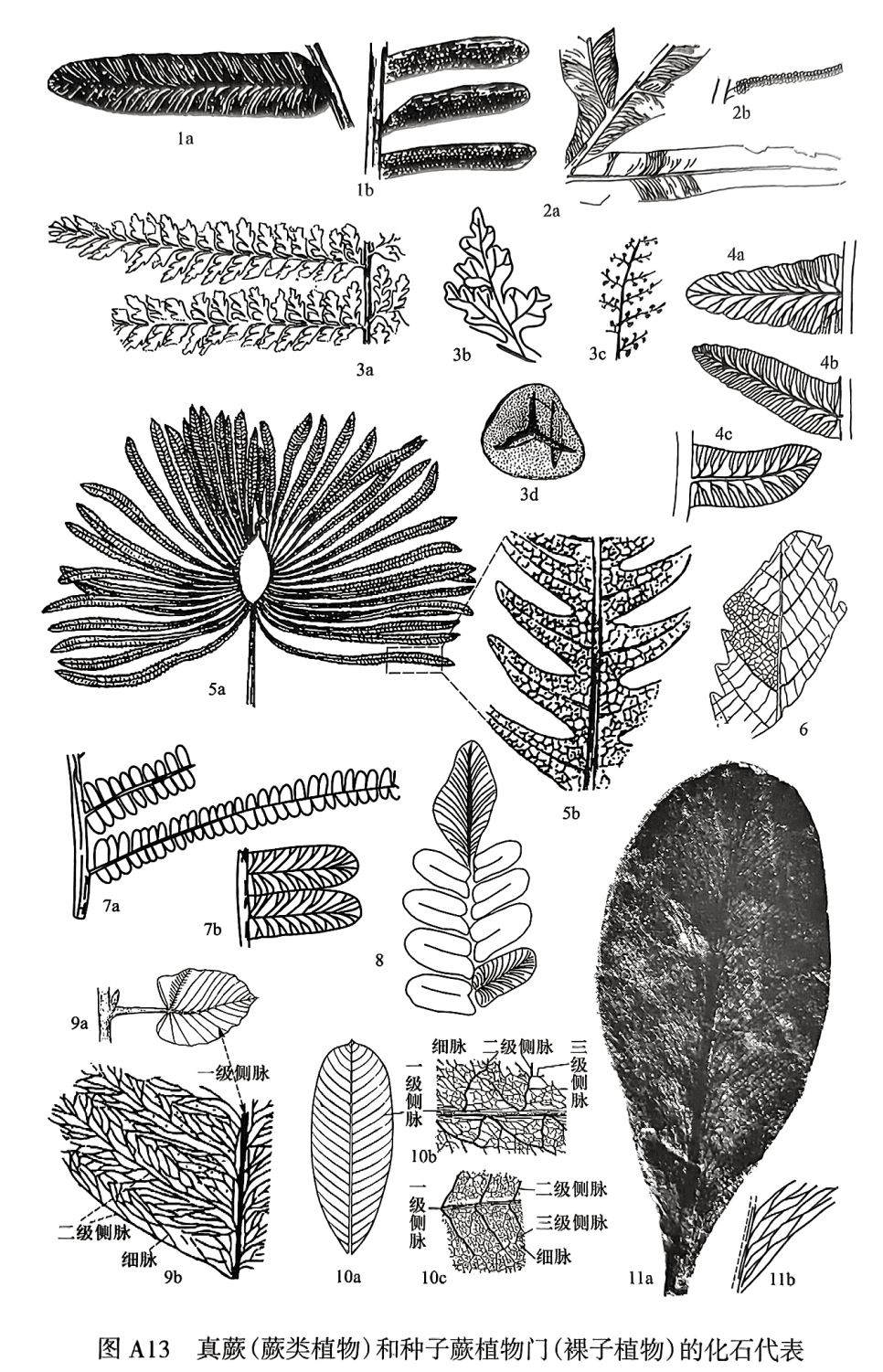
Bernoullia Heer,1877(贝尔瑙蕨):蕨叶1或2次羽状分裂。小羽片长5~6cm,呈线形或剑形,基部收缩,以基部为中心点附着于轴上。中脉粗强,侧脉细密,分叉数次呈束状。实羽片较短,孢子囊成群地排列于侧脉两侧,布满叶的背面(图A13.1a-b)。生活于中三叠世晚期一晚三叠世。
Danaeopsis Heer,1864(拟丹尼蕨):蕨叶大,1~2次羽状分裂;小羽片呈带状,全缘,整个基部着生于羽轴,下延或略收缩。中脉粗,侧脉分叉1~2次,自中脉以锐角伸出,急向外弯,在近叶边缘处二歧式分枝结成稀疏网脉,在基部下延处有邻脉。实羽片同形,孢子囊呈圆形,各自分离,密布于羽片背面,成行排列于侧脉两侧(图A13.2a-b)。生活于晚三叠世,在我国于中三叠世晚期开始出现。
Coniopteris Brongniart,1849(锥叶蕨):蕨叶2~3次羽状分裂,末次羽片以宽角着生于轴上;小羽片为楔羊齿型,基部收缩,边缘分裂为裂片。中脉细弱不显著。有的羽片基部下行具有变态小羽片,它的后瓣裂片呈线形并指向后方。实小羽片的裂片常退化,囊群着生于叶边缘或叶脉末端,具有杯状的囊群盖,在化石印痕上囊群呈扁圆形(图A13.3a-d)。生活于侏罗纪—早白垩世(以侏罗纪最繁盛),广布于世界各地。
Cladophlebis Brongniart,1849(枝脉蕨):蕨叶2~4次羽状分裂,小羽片常较大,且多少呈镰刀形,顶端常为尖或圆凸状,或为全缘或具有齿。羽状脉,侧脉常分叉(图A13.4a—c)。Cladophlebis是一个广泛应用的形态属,一般把自二叠纪至中生代具有上述形态而又未发现生殖羽片的蕨叶都归入本属。大多数的 Cladophlebis可能属于紫其科,其自然属名称为 Todites(似托第蕨)。在三叠纪一早白垩世繁盛。
Dictyophyllum Lindley and Hutton,1834(网叶蕨):蕨叶大,具有长柄,柄顶端二歧式分枝,每个分枝叉轴式分枝向内弯成弧形,外侧辐射状着生羽片。羽片基部相连或不相连,羽片裂成三角形至镰刀状小羽片,各具有中脉,侧脉以直角伸出,结成多角形网格,网内有细脉结成小网(图A13.5a—b,b是a的局部放大),孢子囊群着生于细网孔内。生活于晚三叠世-中侏罗世,分布于北半球。在我国晚三叠世地层中有一个十分常见的 Clathropteris(格脉蕨,晚三叠世-中侏罗世),它与本属十分相似,但格脉蕨的羽片较宽,且分裂较浅,二级侧脉常联结成长方形的大网眼,网眼内又有更小的网格(图A13.6)。
Pecopteris Brongniart,1822(栉羊齿):多次羽状复叶,小羽片大多为两边平行、顶端钝圆的舌形,小羽片全缘。基部全都附着于羽轴,为羽状脉,中脉达顶端。为形态属。大多数为真蕨植物门,少数可能为种子蕨(图A13.7a-b)。生活于石炭纪—二叠纪,分布于世界各地。
2.裸子植物
(1)种子蕨植物门
Neuropteris Brongniart,1822(脉羊齿):奇数或偶数羽状复叶,小羽片为舌形、镰刀形等,基部收缩成心形,以一点附着于羽轴,全缘,顶端尖或钝圆。为羽状脉,中脉明显,伸至小羽片全长之1/2或2/3处就消散,侧脉以狭角分出,多次分叉向外弯(图A13.8)。生活于早石炭世一早二叠世。
Gigantonoclea Koidz(emend. Gu and Zhi,1974)(单网羊齿,属于大羽羊齿类):为羽状复叶或单叶。小羽片(或叶)大,呈披针形、长椭圆形或卵形,边缘为全缘波状或齿状。中脉较粗,侧脉分1~3级,细脉二歧式分枝结成简单网,网眼或长或短,为多角形,具有伴网眼(图A13.9a—b)。生活于早二叠世晚期一晚二叠世,个别残存至早三叠世早期。
Gigantopteris Schenk,1883(大羽羊齿):为单叶,叶大,着生情况不明,整体形状不明。呈倒卵形、纺锤形、歪心形或长椭圆形,边缘为全缘、波状或齿状。叶脉有四级,中脉粗,一级至三级侧脉羽状.三级侧脉联结形成大网眼,并分出细脉,细脉结成小网眼,套叠而成重网状,网眼内有盲脉.中脉上常有邻脉伸出(图A13.10a—c)。生活于晚二叠世(少量残存至早三叠世最早期),为华夏植物群的代表分子。
Glossopteris Brongniart,1828(舌羊齿):为单叶,叶大,呈舌形、披针形或匙形,柄或有或无,边缘为全缘;具有明显的中脉和结成长多边形单网眼的侧脉(图A13.11a—b)。生活于二叠纪—早三叠世,为冈瓦纳植物群的代表分子。
(2)苏铁植物门
苏铁纲(Cycadopsida)
Nilssonia Brongniart,1825(尼尔桑):为羽叶,呈披针形或线形,边缘为全缘或裂成裂片,羽叶基部的叶膜很少分裂。叶膜或裂片着生于羽轴腹面,几乎全部覆盖羽轴。叶脉为简单的平行脉,分叉次数少(图A14.1)。生活于二叠纪-古近纪,主要繁盛于晚三叠世—早白垩世。
本内苏铁纲(Bennettiopsida)
Otozamites Braun,1843(耳羽叶):叶为单叶,羽状。裂片为圆形、宽卵形或披针形,基部收缩成耳状;上耳突出更甚,以基部的一小部分着生于羽轴腹面。裂片互生,上下裂片相挤或叠复。为放射脉(图A14.2)。生活于晚三叠世一早白垩世。
Pterophyllum Brongniart,1824(侧羽叶):叶为单叶,羽状。裂片基部全部附着于羽轴的两侧,为线形、扁针形或舌形,两侧边平行。为平行脉,分叉1~3次(图A14.3)。生活于晚石炭世一早白垩世,在晚三叠世一侏罗纪最盛。
Ptilophyllum Morris,1840(毛羽叶):叶为单叶,羽状。裂片线形,似侧羽叶,但基部全部着生于羽轴腹面,斜伸。裂片基部上边收缩成圆形,下边略向下延;上面的裂片常被下面裂片叠覆一部分。叶脉平行或呈放射状(图A14.4a-b)。生活于晚三叠世—白垩纪。
(3)银杏植物门
银杏目(Ginkgoales)
Ginkgoites Seward,1919(emend. Florin,1936)(似银杏):叶形与现代银杏相近。具有长柄,呈扇形、肾形或楔形。为扇状脉。叶常二歧式分裂成2~8个或更多个最后裂片,每个裂片内有平行脉4~6条或更多。叶表皮细胞结构与现代银杏有差别(图A14.5)。生活于早二叠世一新近纪,在侏罗纪一早白垩世最盛。
茨康目(Czekanowskiales)
Phoenicopsis Heer,1877(拟刺葵):叶线形,无柄,不分裂;常6~20枚簇生于短枝上;短枝基部布满披鳞片状小叶,线形叶宽0.2~2cm,一般宽4~10mm,长可达20mm,宽度变化不大。平行脉偶有分叉(图A14.6)。生活于晚三叠世晚期一晚白垩世,极盛于早、中侏罗世。
(4)松柏植物门
科达纲(Cordaitopsida)
Cordaites Unger,1850(科达):为细高乔木,叶子密集螺旋着生于顶端小枝。叶呈带形,边缘为全缘,无柄。为平行脉,或具有脉间纹或否(图A14.7)。生活于石炭纪一二叠纪,分布于欧美及中国热带植物地理区。
松柏纲(Coniferopsida)
Podozamites Braun,1843(苏铁杉):枝轴细;叶稀,螺旋状着生,呈假两列状;呈椭圆形、披针形或线形。叶脉细,平行叶边缘,至顶端常聚缩(图A14.8)。生活于晚三叠世一早白垩世,分布于北半球。
Ullmannia Goeppert,1825(鳞杉):乔木,小枝排列不规则;叶呈卵形、短披针形或披针形,基部宽而下延。为单脉,叶表面具有细纵纹,呈覆瓦螺旋状排列(图A14.9)。生活于晚二叠世,分布于欧洲、亚洲。
3.被子植物
Magnolia Linn,1753(木兰):为落叶乔木。为大型单叶,呈卵形至阔椭圆形,顶端具有短尖头.基部呈宽楔形至钝圆形。中脉粗强,羽状侧脉以宽角伸出,至近叶缘向上弯,彼此联结成脉环.细脉多呈角网状(图A14.10)。生活于晚白垩世—现代,在古近纪时广布于北半球,在现代仅分布于北美、东亚。
Salix Linn,1753(柳):为落叶乔木。为单叶,呈披针形至线状披针形,顶端渐尖,基部呈宽楔形.边缘或具有细锯齿或为全缘。中脉明显而粗强;羽状侧脉甚纤细,至叶边缘向前弯曲,彼此联结;细脉呈网状(图A14.11)。生活于晚白垩世-现代,在温带植物区常见。
