第一节 前寒武纪的生物界
一、太古宙一中元古代生命记录
目前最早的生命记录发现于3500Ma之前的沉积岩中,包括格陵兰岛3800Ma的地层中发现的叠层石、生物成因的矿物和地球化学证据(Mojzsis et al.,1996; Nutman et al.,2016),加拿大3770Ma的玉髓里面的丝状化石(Dodd et al.,2017),西澳大利亚3400~3500Ma的 Warrawoona群地层中的丝状和链状细胞体(Schopf et al.,1994)等。自3500Ma生命出现之后的长达10亿年的时间里,原核生物几乎是地球上唯一的生命存在形式。它们大多属于厌氧化能自养细菌,这个时期的生态系统是建立在化能自养基础上的微生物生态系统。
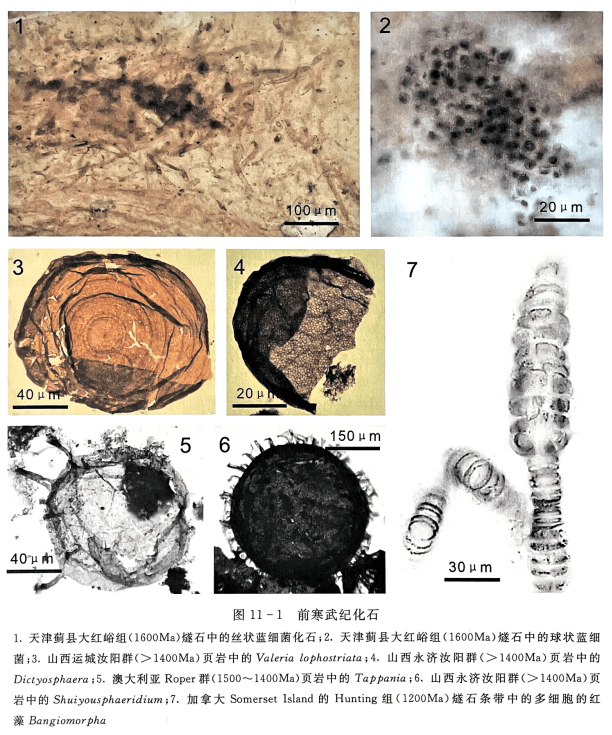
从古元古代开始蓝细菌开始取代化能自养微生物,逐渐成为生态系统的主要组成部分。进入中元古代,蓝细菌大量繁盛,在世界范围内沉积了大规模的碳酸盐岩沉积,这意味着以蓝细菌类为主的沉积碳酸盐和建造叠层石的微生物广泛分布和繁盛于陆缘浅海和滨海环境。在元古宙长达20亿年的时间里,蓝细菌一直是生物圈主要的(占优势)的生物类群,也是全球生态系统中最重要的初级生产者。蓝细菌化石被广泛地报道于世界各地的地层中,包括加拿大1900Ma的 Gunflint铁建造(Awramik,1978),中国华北1600~1400Ma的蓟县群(Zhang,1985; Lee and Golubic,1998; Shi et al.,2017),西伯利亚1500Ma的 Kotuikan组(Vorob’ eva et al.,2015),加拿大1200Ma的 Dismal Lakes群(Horodyski and Donaldson,1980)等。
地球上最早的真核生物是报道于1700Ma以后的地层中的具有抗酸碱的有机质壁化石,包括外壁同心纹纹饰的 Valeria lophostriata,具有六边形纹饰的 Dictyosphaera 和 Shuiyousphaeridium,具有凸起的 Tappania 和 Shuiyousphaeridium(Adam,2014;Agié et al.,2015; Pang et al.,2015; Javaux et al.,2001; Nagovitsin,2009; Peng et al.,2009; Yin et al.,2005)。 距今1200Ma开始,加拿大 Somer-set Island的 Hunting 组燧石条带中出现了最早的多细胞的红藻 Bangiomorpha(Butterfield,2000)(图11-1)。
二、新元古代的生物界
新元古代是地球生命演化的重要时期,在地球早期生物演化的过程中,受太古宙末期和新元古代后期两次大氧化事件的影响,新元古代后期地球的大气氧含量有了显著提升,接近于现今大气氧含量。地球上的海洋化学条件也有了利于生物进化的改观。新元古代开始出现一系列复杂真核生物的记录,为寒武纪初期生命大爆发打下了坚实基础。按时间顺序,新元古代的生物记录主要包括宋洛生物群、蓝田生物群、瓮安生物群、庙河生物群、埃迪卡拉生物群等。
1.宋洛生物群
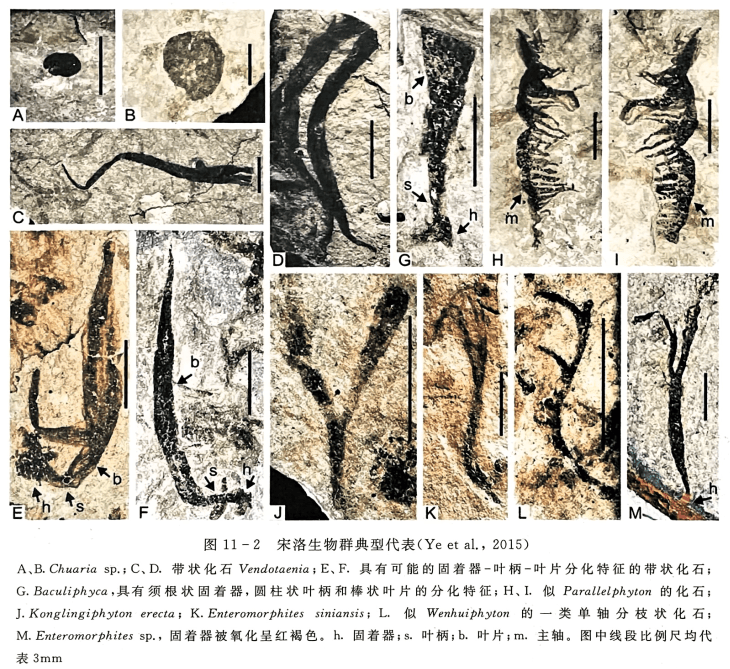
宋洛生物群是指发现于神农架东部地区宋洛乡,成冰纪南沱组冰碛岩夹层的黑色页岩、粉砂质泥岩中的,以宏体藻类为代表的生物组合。20世纪八九十年代,朱士兴、瞿乐生等老一辈地质工作者在神农架宋洛地区工作时,在成冰纪南沱组杂砾岩的碳质页岩夹层中,发现了少量带状碳质压膜化石 Vendot-aenia sp.和 Tyrasotaenia ef. podolica,并将它们解释为大型褐藻化石(李铨和冷坚,1991)。 但由于化石数量较少,且多为碎片,并未引起关注。2012年以来,经童金南、叶琴等再次发掘和研究,发现该地区的碳质化石不仅具有宏观体积,而且大部分表现出明显的形态分异,其中既包括一些形态简单、延续时间较长的化石类型,同时也包括一些形态复杂、被解释为底栖固着生活的宏体藻类化石,因此认为它们主要是一个以多细胞宏体藻类为主的化石组合(Ye et al.,2015)。
通过研究,宋洛生物群化石面貌至少包括10余个不同形态的类群,其中具代表性的形态类型有:①圆盘形或椭圆形(图11-2A、B),其直径通常介于2~10mm之间,平均5.5mm,以 Chuaria 为代表。②带状化石(图11-2C、D),化石呈两侧平行,两端逐渐尖缩的带状体,无分枝,常扭折,折叠或弯曲状产出,宽0.1~2mm,长度不一,最长可达20mm,常被认为是 Vendotaenia 属或其相关类群。在这类带状化石中,少量化石表现为不仅具有叶片、叶柄,基部还有须根状固着器(图11-2E、F)。③棒状类型(图11-2G),这类化石高达7mm,最宽处为2mm。其最大特征是具有须根状固着器、棒状叶柄和叶片的分化特征,可归属为 Baculiphyca。④可能的分枝类型,包括 Konglingiphyton erecta(图11-2J,一次分枝,分枝向上变宽,分枝宽度由下部的0.1~0.3mm向上变宽至 0.8mm,分枝角约 30°)、Enteromorphites siniansis(图11-2K,等二歧式分枝或者分枝向上略有变窄,分枝宽约0.2mm)、En-teromorphites sp.(图11-2M,与E. siniansis相似,但其下部具有一个膨胀的叶柄和一个球形的固着器)。⑤假单轴式分枝类型,又包括两个属种化石。其一特征是稀疏的圆柱状分枝从主轴一侧伸出,主轴宽0.17mm,长5.1mm,侧枝宽0.06~0.15mm,长0.8~1.2mm,分枝间距为0.8~1.1mm,分枝角较大,可达60°~90°(图11-2L)。这类分枝方式与埃迪卡拉纪翁会生物群中的 Wenghuiphyton erecta相似,不同点在于后者侧枝有二歧分枝现象。其二是从主轴一侧分散出大量侧枝,且侧枝向远端有变细变尖现象。主轴最宽处约1mm,长达15mm,侧枝宽0.2~0.6mm,长1.5~3.1mm,分枝角近垂直,达到85°(图11-2H、I)。此化石类型与中寒武世凯里生物群中的一类可能的藻类化石 Parallelphytontipica 形态特征相似,但后者的侧枝也存在向远端二歧分枝的现象。
宋洛生物群的发现对于认识“雪球地球”时期的环境特征、生物面貌和两者之间的关系,以及早期宏体藻类形态的演替,都是全新的资料或重要的补充(Corsetti,2015)。在以往的认知中,全球成冰纪由于低温与冰冻的极端环境制约了生物的发展,古生物化石的报道甚为贫乏,仅少数地方的间冰期沉积中有部分微生物化石的报道。但宋洛生物群的发现表明,即使在典型的冰期沉积南沱组中至少已发现了多种类型的宏体藻类化石,部分化石还表现为具固着器、分枝的特征,充分说明了宏体藻类在成冰纪已具有一定的分化特征,同时具有一定的多样性和复杂性,弥补了世界范围内的成冰纪宏体藻类化石资料的极度不足(空白)。另外,值得注意的是,上述宋洛生物群不仅包括一些形态简单、延续时间较长的化石类型(如 Chuaria 和 Vendotaenia),而且也包括一些形态复杂、被解释为底栖固着生活的宏体藻类化石(如 Baculiphyca sp.、Konglingiphyton erecta、Enteromorphites siniansis等)。这不仅说明以南沱组为代表的冰川作用并没有使生物群完全灭绝,以蓝田生物群为代表的埃迪卡拉纪后生生物也并不是随着埃迪卡拉纪的开始而突然爆发的,而是在以南沱冰碛岩为代表的成冰纪生物群的基础上发展而来的,因此极端环境条件可能是造就新类型生命的关键因素。而且,不同于现代冰川中的微体生物具有较为广泛的生活范围,宋洛南沱组泥质页岩夹层中的底栖宏体藻类所代表的环境条件可能表明,它们不但需要较为稳定的生存空间,同时它们纤细脆弱的须根状或球状固着器不适于生活在坚硬的石质底质,可能更适于生活在泥质底质上。此外,藻类的生长、繁殖需要依赖阳光进行光合作用,因而其生活环境应是开放水域的透光带。因此,宋洛生物群的发现说明在 Marinoan冰期(南沱冰期),至少在中纬度的华南滨岸环境存在开放水域,存在适合宏体底栖藻类生存的底质,为理解“雪球地球”的强度和范围提供了重要的化石证据(Ye et al.,2015)。
2. 蓝田生物群
蓝田生物群产于安徽省黄山市休宁县蓝田地区,化石主要以碳质压膜的形式保存在埃迪卡拉系下部蓝田组二段的黑色页岩中。该生物群最早由毕治国、王贤方于1981年发现,并由邢裕盛等(1985)正式报道,初步描述了5属。此后不同学者相继对该地区宏体化石进行大量挖掘和研究,随着化石面貌的不断增加,闫永奎等(1992)将该化石组合命名为“蓝田植物群”。蓝田生物群先后被描述的化石种多达50个以上,后经袁训来等系统修订,归入12~15个种一级的分类单元(Yuan et al.,1999),基本为碳质压膜保存的宏体藻类化石,没有发现可靠的动物化石。近年来,袁训来团队对该生物群开展了更加详细的系统研究,有关这一化石生物群的总体面貌和时代等方面的研究取得了新进展。在化石面貌方面,现已发现24种不同类型的宏体化石,按照生物属性可以划分为宏体藻类、后生动物和疑难化石三大类群(图11-3),分别是:宏体藻类 Anhuiphyton lineatum、Chuaria spp.、Doushantuophyton lineare、Doushantuophyton rigidulum、Doushantuophyton cometa、 Enteromorphites siniansis、Flabellophyton lantianensis、Flabellophyton spp.、Grypania spiralis、Huangshanophyton fluticulo-sum、Marpolia spissa;后生动物 Lantianella laevis、Lantianella annularis、Piyuania cyathiformis、Qianchuania fusiformis、Xiuningella rara;疑难化石 Orbisiana linearis、Unnamed Form A、UnnamedForm B、Unnamed Form C、Unnamed Form D(袁训来等,2016; Wan et al.,2016)。该生物群不仅包含了形态多样的宏体藻类,也有具触手和类似肠道特征、形态可与现代腔肠动物或蠕虫类相比较的后生动物,因此正式更名为“蓝田生物群”(袁训来等,2012)。在时代方面,根据化学地层学、沉积序列并结合区域地层对比,认为蓝田生物群属于埃迪卡拉纪早期的宏体多细胞生物群,早于以往报道的所有埃迪卡拉生物群,时代限定在635~580Ma之间(Yuan et al.,2011)。
蓝田生物群为我们重新认识早期复杂宏体生命打开了一个新的窗口(Narbonne,2011)。一方面,它是地球早期形态简单的微体真核生物向体型结构复杂的多细胞宏体真核生物演化的重要环节,显示了在新元古代雪球地球事件刚刚结束后不久,形态多样化的宏体真核生物,包括海藻和后生动物,就发生了快速辐射(Yuan etal.,2011);另一方面,从蓝田生物群的总体特征及相关环境信息推断,它生活在水深100~200m之间的静水、有氧的海洋环境中,预示着多细胞宏体生物的起源和早期演化很可能发生在较深水的安静环境中,并在埃迪卡拉纪中晚期逐步迁移和扩散到较浅水的近岸环境中(袁训来等,2016)。

3.瓮安生物群
瓮安生物群产自贵州省瓮安县瓮安磷矿埃迪卡拉系陡山沱组中上部磷块岩之中,距今约609Ma(Zhou et al.,2017a)。瓮安生物群的研究历史要追溯到20世纪80年代初期,1984年,朱士兴等描述了瓮安磷矿中的一种多细胞藻类(朱士兴等,1984),随后陈孟莪和刘魁梧首次报道了瓮安磷矿陡山沱组磷块岩中的大型球状化石和疑源类(陈孟莪和刘魁梧,1986),揭开了瓮安生物群的研究序幕。此后,瓮安陡山沱组中一系列精美保存的化石被发现,并于1993年被命名,它是指由底栖的多细胞藻类、浮游疑源类、丝状和球状蓝藻组成的晚前寒武纪生物群(袁训来等,1993)。1998年,国际顶级期刊 Nature 和Science 几乎同时报道了瓮安生物群中的“后生动物胚胎化石”(Xiao et al.,1998a)和“具细胞结构的海绵动物及其胚胎化石”(Li etal.,1998),将多细胞动物出现的化石记录从寒武纪向前大大推进,使得瓮安生物群受到国际学术界的极大关注,迅速成为早期生命研究的前沿热点。
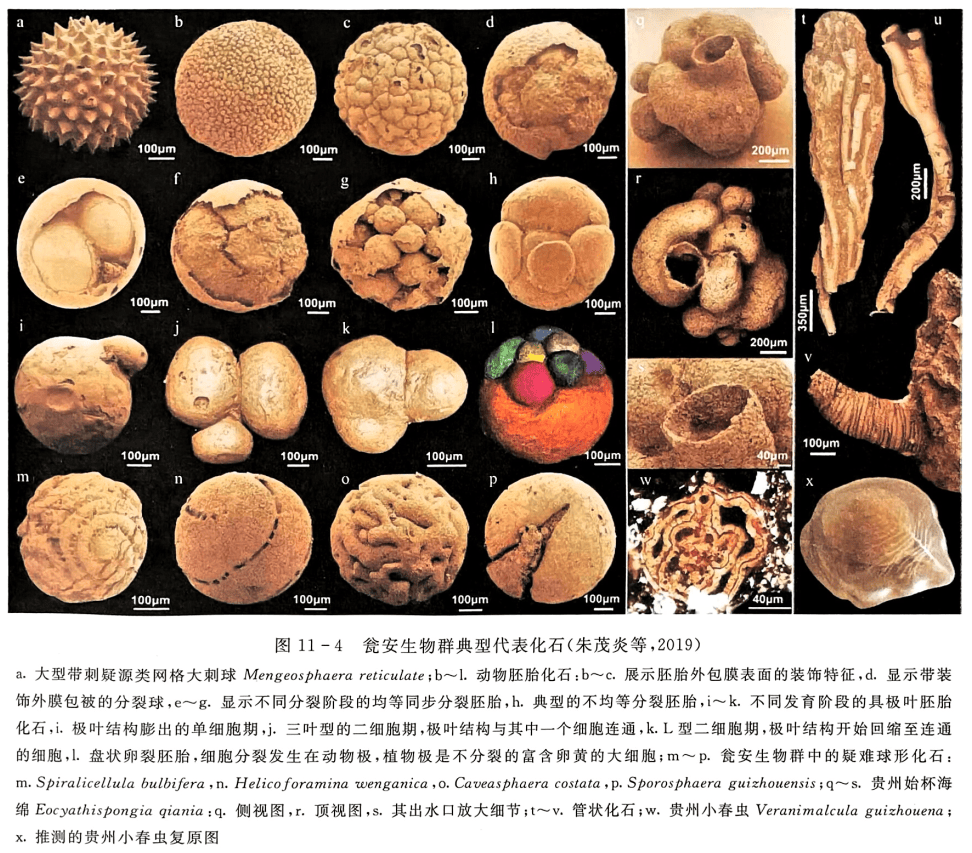
到目前为止,瓮安生物群所涉及的生物内涵和所包含的属种发生了很大变化,使瓮安生物群不仅包含丝状和球状蓝藻类、具组织结构和分化的多细胞藻类和大型具刺疑源类等化石,还包含丰富的后生动物休眠卵和胚胎化石以及少量可疑的早期后生动物微型成体化石(Xiaoetal.,2014a)。根据对已发表资料的统计,瓮安生物群已描述的上述各类化石达51属86种(图11-4,尹崇玉等,2007;Xiao et al.,2014b及其内参考文献),其中丝状及球状蓝藻7属12种(含3个未定种) Archaeophycus yunnanensis、Obruchevella parva、Salome nunavutensis、Salome hubeiensis、Siphonophycus septatum、Siphono-phycus robustum、Siphonophycus typicum、Siphonophycus kestron、Siphonophycus solidum、Eozy-gion sp.、Gloeodiniopsis sp.、Myxococoides sp.;多细胞藻类6属8种 Gremiphyca corymbiata、Par-am ecia incognata、Sarcinophycus radiatus、Thallophyca ramosa、Thallophyca corrugata、Thallo-phycoides phloeatus、Wengania globosa、Wengania exquisita;疑源类和动物胚胎化石31属56种(含未定种2个) Appendisphaera grandis、Appendisphaera tenuis、Asterocapsoides robustus、Astero-capsoides sinensis、Asterocapsoides wenganensis、Bacatisphaera baokangensis、Baltisphaeridium ri-gidium、Bullatosphaera sp.、Caveasphaera costate、 Cavaspina acuminate、 Cavaspina basiconica、Cymatiosphaeroides yinii、Dicrospinasphaera virgate、Dicrospinasphaera zhangii、Distosphaera speciose、Eotylotopalla dactylos、 Eotylotopalla delicate、Ericiasphaera magna、 Ericiasphaera rigida、 Helicoforamina wenganica、 Hocosphaeridium scaberfacium、 Hocosphaeridium anozos、Knollisphaeridium? bifurcatum、Knollisphaeridium ef. gravestockii、Knollisphaeridium maximum、 Knollisphaeridium triangulum、 Megasphaera inornate、 Megasphaera cymbala、Megasphaera ornate、Megasphaera patella、Megasphaera puncticulosa、Meghystrichosphaeridium magnificum、Mengeosphaera chadianensis、Mengeosphaera eccentrica、Mengeosphaera reticulata、Mengeosphaera sp.、 Papillomembrana boletiformis、 Papillomembrana compta、 Pustulisphaera membranacea、 Sinosphaera speciosa、 Sinosphaera variabilis、 Spiralicellula bulbifera、Sporosphaera guizhouensis、 Taedigerasphaera lappacea、 Taeniosphaera doushantuoensis、Tanarium conoideum、 Tanarium digitiforme、 Tanarium victor、 Tianzhushania polysiphonia、Tianzhushania rara、Tianzhushania spinosa、Variomargosphaeridium gracile、Variomargospha-eridium litoschum、Vulcanisphaera phacelosa、Weissiella brevis、Yinitianzhushania tuberifera;可能的后生动物化石 7 属 10 种 Crassitubus costatus、 Eocyathispongia qiania、 Quadeatitubus orbigoniatus、Ramitubus increscens、Ramitubus decrescens、Sinocyclocyclicus guizhouensis、Sinocy-clocyclicus centriporatus、Sinoquadraticus poratus、Sinoquadraticus wenganensis、Vernanimalcula guizhouena。
其中动物胚胎化石作为迄今最古老的后生动物化石之一,一直以来是瓮安生物群中最具影响力也是最有争议的化石。这些化石呈球形,大小在200~800μm之间,壳体表面光滑或具板状、瘤状、脑纹状纹饰,壳体内部分别包裹着1个、2个、4个、8个、16个乃至2°个有规律出现的小球,且具有不同数量小球的球状化石个体大小也大致相同。这类化石最初由薛耀松等(1995)发现,将之与现生绿藻门下的团藻进行对比。1998年,肖书海等认为这些球状化石具有等体积细胞分裂的特征,即细胞总体积不变,单个细胞体积随细胞分裂次数增多(按2的指数形式增长,即细胞个数从2°到2¹到2²⋯⋯,直到成百上千个细胞)而递减。这些特征正是后生动物胚胎早期卵裂的特征,据此将内含2、4、8等多个细胞的化石解释为处于卵裂阶段的动物胚胎,将内含单个细胞且有表面装饰包壳的球状微体化石解释为处于休眠状态的合子,并认为可与现代鳃足类节肢动物休眠卵进行形态学上的对比(Xiaoetal.,1998a)。后来的研究还发现更多不同类型的胚胎化石(朱茂炎等,2019),并观察到这些球状化石内部细胞分裂到数百个之后,出现了营养细胞和繁殖细胞的分化,预示其发育过程发生过程序性细胞凋亡,因而有可能是某种基干类群动物,但也不能排除是某种多细胞真核藻类(Chen etal.,2014)。目前有关这些球状化石的亲缘关系仍争议不断(Cunningham et al.,2017),未来的研究需要更多的证据才能作出更为明确的解释。
瓮安生物群中可能的后生动物成体化石同样引人注目,包括海绵动物化石、微管状化石和两侧对称动物化石等,均遭受一定程度的质疑(Xiao et al.,2014a;朱茂炎等,2019及其内参考文献)。2015年,殷宗军等在瓮安生物群中发现了一枚海绵动物实体标本(图11-4q~s),这一化石十分微小,体积只有2~3mm³,但保存了精美的细胞结构和完好的水沟系统(Yin et al.,2015),是迄今为止全球发现的最古老的可靠海绵化石的记录,该发现不仅将海绵动物在地球上出现的实证记录从寒武纪向前推进了6千万年,还意味着复杂的多细胞动物起源的时间可能远远早于古生物学家的传统推测。
此外,瓮安生物群中常见的多细胞藻类,细胞和组织结构清晰可见,甚至部分标本可见细胞壁上的微细构造,为研究其内部结构、发育特征及亲缘关系等提供了重要的科学依据(袁训来等,2002);对于大型具刺疑源类化石来说,虽然亲缘关系及生物分类位置尚难以确定,但它们结构精美、类型多样,具有明显时代特征,是该时期海洋浮游生态系统的重要角色,代表了该时期浮游真核生物的演化水平,同时也是埃迪卡拉系内部划分和区域/洲际对比的重要标志。
总之,瓮安生物群这一磷酸盐化特异埋藏化石库以三维立体的形式保存了大量非矿化生物的细胞和亚细胞结构,透过瓮安生物群这一独特窗口,让我们有机会在细胞和组织水平上研究6亿年前生物的进化。
4.庙河生物群
1984年,朱为庆和陈孟莪首次报道了湖北秭归县庙河地区陡山沱组-灯影组过渡地层(庙河段)黑色页岩中的一类宏体碳质压膜化石(朱为庆和陈孟莪,1984)。1990年底,陈孟莪和肖宗正重新奔赴该地,采集了40余块宏体藻类和其他可能的动物化石标本,并将该地区产出的这一特殊化石组合命名为“庙河生物群”(陈孟莪和肖宗正,1991)。随后,不同课题组在这个化石群中发现了更多化石,丁莲芳等(1996)在《震旦纪庙河生物群》一书中描述了140多个种。Xiao等(2002)在大量古生物学研究工作以及前人研究的基础上,将庙河生物群内的化石进行重新厘定,共描述并甄别出17属20种(图11-5):Aggregatosphaera miaoheensis、Anomalophyton zhangzhongyingi、Baculiphyca taeniata、Belta-nelliformis brunsae、Calyptrina striata、Cucullus fraudulentus、Doushantuophyton lineare、Doush-antuophyton quyuani、Enteromorphites siniansis、Glomulus filamentum、Jiuqunaoella simplicis、Konglingiphyton erecta、Liulingjitaenia alloplecta、Longifunciculum dissolutum、Miaohephytonbifurcatum、 Protoconites minor、 Sinocylindra yunnanensis、 Sinospongia chenjunyuani、Sinospongia typica 和 Siphonophycus solidum。并认为其中大部分可能与现生的三大高级藻类分支——绿藻、红藻、褐藻有着亲缘关系,属真核多细胞藻类,同时至少5种类别可能是原始的后生动物化石。例如庙河生物群中最普遍的双叉庙河藻(Miaohephyton bifurcatum),个体呈“Y”字形,表面可见瘤状结构,解释为生殖窝,与现生褐藻门的墨角藻属亲缘相近,可能是早期褐藻起源的重要证据(Xiaoetal.,1998b);而环纹杯状管(Calyptrina striata)具有加厚的环带,形态上类似某些现生腔肠动物的栖居管(Xiao et al.,2002);此外,八臂仙母虫(Eoandromeda octobrachiata)是一类体型奇特的球囊状宏体化石,由8个完全相同的旋臂组成,曾被认为是遗迹化石(丁莲芳等,1996),后认为其与八射珊瑚或栉水母动物有关,可能是刺胞动物的祖先类型(Zhu et al.,2008)。
庙河生物群在湖北峡东地区的分布范围比较局限,三十年以来,除庙河地区以外,没有在其他剖面中发现,因此对庙河生物群的研究一度陷入了沉寂。直到近几年,童金南、安志辉、叶琴等在神农架一宜昌地区埃迪卡拉纪晚期庙河段黑色页岩中新发现多个宏体化石产地,如芝麻坪、麻溪和三里荒等,从而扩大了庙河生物群的地理分布范围,为埃迪卡拉纪多细胞生物群研究提供了新的参考资料(An etal.,2015;Ye et al.,2019)。同时,伴随新的化石产出地点的发现,有关庙河生物群的赋存层位(即庙河段)的不同认识(An et al.,2015; Zhou et al.,2017b)也使得该生物群的时代归属产生了很大的争议。 到目前为止,对庙河段的地层对比问题主要有两种不同的对比方案。传统上人们将庙河地区两套页岩夹一套白云岩的组合总体与九龙湾剖面陡山沱组第四段进行对比(Zhouetal.,2017b),而安志辉等通过大量野外调查,综合古生物化石、层序地层和化学地层的研究,认为庙河段要晚于陡山沱组第四段沉积,应当与灯影组石板滩段下部进行对比(An et al.,2015)。目前庙河生物群的最小年龄被限制在551Ma(Condonetal.,2005),如能在陡山沱组第四段和/或灯影组石板滩段下部开展同位素测年工作,将是解决上述地层对比争议的重要方法之一。
庙河生物群是以底栖固着藻类为主的复杂宏体生物群,是宏体藻类高度繁盛的时期。与新元古代之前的宏体藻类相比,该时期的宏体藻类形态稳定,具高的丰度和分异度,分枝清楚,器官分化明显,具有现生藻类的诸多形态和结构特点,代表了宏体藻类演化史上的一次辐射性事件,是了解寒武纪生命大爆发前夕生物多细胞化、组织化和生物宏体形态多样化的重要窗口之一。

5.埃迪卡拉生物群
埃迪卡拉生物群最早由 Sprigg于1946年发现于澳大利亚南部埃迪卡拉山前寒武纪末期5.5亿年前的庞德石英砂岩中,目前几乎在全球都有发现(Boag et al.,2016; Waggnoer,2003)。它主要分为3个生物组合(Narbonne,2005; Waggnoer,2003),自下而上分别为:阿瓦隆生物群组合(the Avalon as-semblage,575~560 Ma),主要包括典型的叶状化石 arboreomorphs和 rangeomorphs分子,代表地为纽芬兰和英格兰地区,属于深水边缘斜坡和盆地相环境;白海生物群组合(the White Sea assemblage,560~550 Ma),数量最为丰富,主要包括 dickinsoniomorphs、bilateralomorphs、triradialomorphs/ tri-brachiomorphs等(图11-6),代表地点为南澳大利亚弗林德斯山脉和俄罗斯白海等地,分布在近海环境;纳玛生物群组合(the Nama assemblage,550~541Ma),由 erniettomorph(图11-6g~i)、rangeo-morph 和少量具有矿化骨骼动物如 Cloudina 和 Namacalathus 组成,代表地为纳米比亚、华南、加拿大和英国西部地区,是近岸滨岸环境。这3个埃迪卡拉纪生物群组合见证了埃迪卡拉纪生物的形态空间演化过程,即埃迪卡拉生物体的形态空间在阿瓦隆生物组合即已达到最大程度,并在之后的白海生物组合和纳玛生物组合中基本保持不变,尽管在生物类型丰度方面,白海生物组合远高于阿瓦隆和纳玛生物组合。埃迪卡拉生物群中这种生物体形态演化与生物类型丰度之间的脱耦性与寒武纪生物大爆发类似,可能代表着二者在生物演化模式上具有相同的驱动机制(Shen et al.,2008; Xiao and Laflamme,2009)。

埃迪卡拉生物群是寒武纪生物大爆发前夕最引人注目的复杂生物群,该生物群从一开始被发现便解释为与寒武纪之后和现代海洋中的动物相关的动物祖先类群。但是埃迪卡拉生物群中的化石个体较大,形态多样且奇特,呈软躯体保存,缺乏矿化外壳和骨骼,几乎都不能与寒武纪之后乃至现今的生物进行很好的形态对比,因此有关埃迪卡拉生物属性和生活方式存在各种各样不同的假说,如真菌、地衣、原核生物、原始多细胞动物等。1992年,Seilacher提出一个新的生物门类———文德动物门(Vendozoa)或文德生物群(Vendobionta),以示埃迪卡拉生物群在生命演化史中的特殊性。他认为埃迪卡拉生物群中即使有可归人现生的门甚至动物界的化石,那也只是少数,大部分类型是一群在地质历史上“昙花一现”、已经完全灭绝了的生物,并认为这些化石是由单细胞通过特定的方式构成的,它们的演化是以表面积的扩大为特征,是生物演化史上一次失败的尝试(Seilacher,1989,1992)。另一部分学者认为埃迪卡拉生物群代表了寒武纪大爆发前夕出现的大量存活时间很短的动物祖先分子,可能属于动物系统树底部不同位置上的干群支系(DunnandLiu,2019)。支持的证据包括具有肌肉收缩运动和动物标志化合物的 Dickinsonia、可能的两侧对称动物如 Kimberella 和 Parvancarina、刺细胞动物 Haootia 和Bjarmia、海绵 Thectardis 和 Coronacollina(Muscente et al.,2018及其内参考文献)。近年来,在华南三峡地区灯影组石板滩段灰岩中发现了多种类型的埃迪卡拉生物群典型分子,还包括一些复杂两侧对称动物实体化石和遗迹化石,如三叶形蠕形动物实体与其运动痕迹一起保存的化石(图11-6j)和具有成对附肢动物的爬行觅食迹化石等,为埃迪卡拉纪晚期两侧对称动物干群的存在提供了可靠的化石证据(Chen et al.,2018,2019),是对寒武纪大爆发幕式演化过程的最有力支持(朱茂炎等,2019)。