5 腕足动物
内容节选自《古生物学》(童金南主编,2021年版)第五章 古无脊椎动物 第五节 腕足动物
腕足动物亦称腕足类(brachipods),是海生、底栖、单体(无群体)群居生活的真体腔(不分节)且两侧对称的无脊椎动物门类。体外披着两瓣大小不等的壳,壳质主要为钙质或几丁磷灰质。腕足动物是滤食性生物,其摄食器官是纤毛腕。腕足动物在地史时期相当繁盛,自寒武纪至第四纪均有化石记录。据统计,已描述的腕足动物有近4220属(Williams, et al.,2007),种数估计超过50000种。然而,现存腕足动物仅约100属300余种。腕足动物化石在确定地质时代和古环境恢复等方面具有重要意义。
一、腕足动物的软体特征
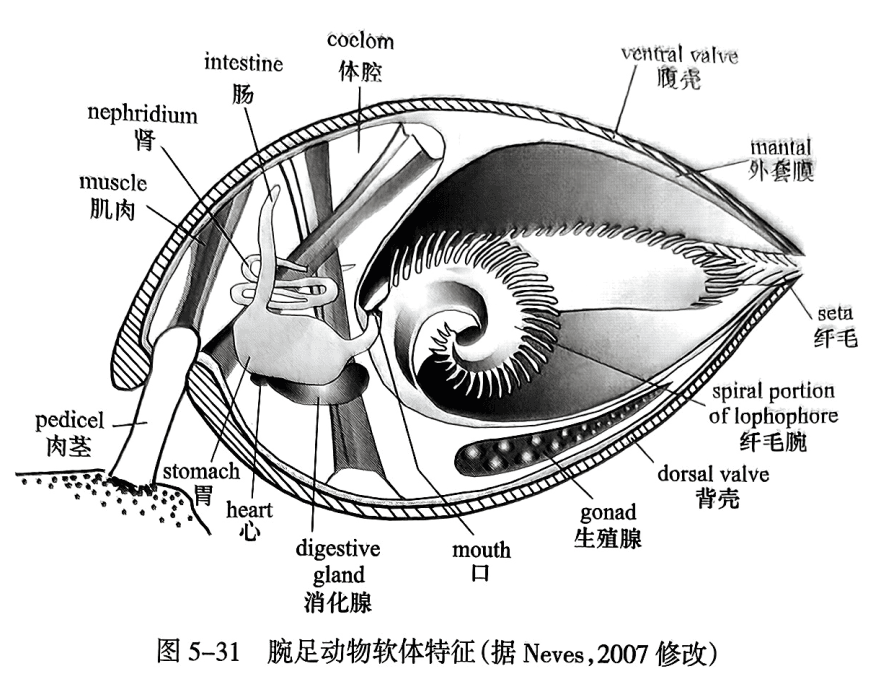
腕足动物软体的最外部是外套膜,紧贴在两枚硬壳的内面,围成外套腔,被中间的横隔膜分成内脏腔和腕腔两部分。内脏腔又名体腔,位于身体后部,约占身体的三分之一;腕腔是容纳纤毛腕的空间,位于前部(图5-31)。软体大部分集中在内脏腔内,包括:
(1)消化排泄器官。口位于两条纤毛腕的着生点之间,有横隔膜连接食道、胃和肠。除舌形贝类外,其他各类无肛门,粪粒从口排出。有一对肾管,一端通入腕腔,兼司生殖管的作用。
(2)神经系统。围绕食道的神经环是神经中枢,神经延伸到纤毛腕和外套膜上。
(3)循环系统。血管能收缩的部分构成心脏,血管在内脏腔中开口。血液无色,无细胞,可凝固。
(4)生殖腺。雌雄异体,少数雌雄同体。常有两对生殖腺囊,背方和腹方各一对。生殖腺在化石上的痕迹,称生殖腺痕(genital marking)。
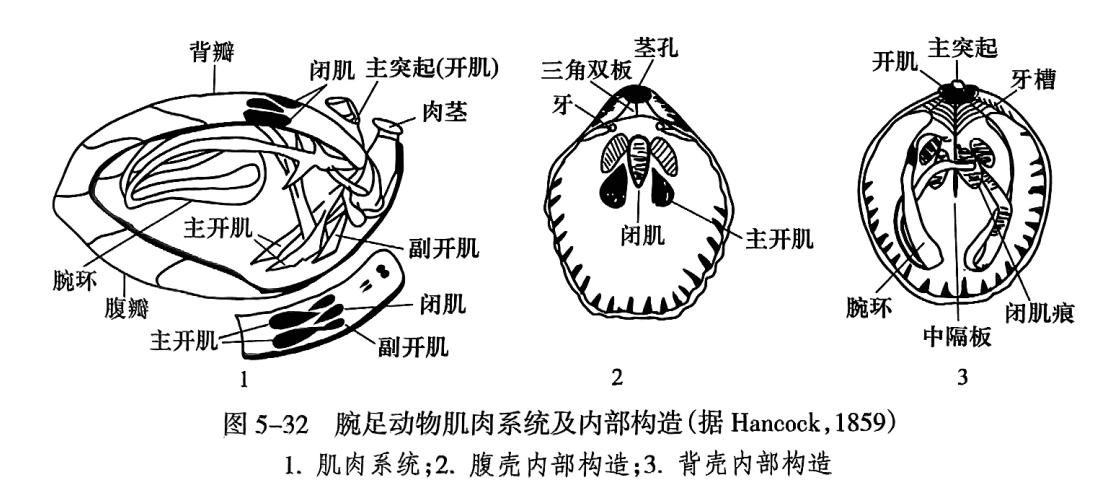
(5)肌肉系统。司壳的开闭及肉茎(pedicle)的伸缩和扭转(图5-32),包括闭肌、开肌、肉茎调整肌。闭肌(adductor)有一对,位于腹壳内脏腔区中部;向背方分成两对,附着于背壳内脏腔区中部。主开肌(diductor)有一对,一端附着于腹壳闭肌的前侧方,另一端附着于背壳的主突起上。其他有5个肉茎调整肌和副开肌等。无铰类的肌肉系统更为复杂,除司壳开闭及肉茎伸缩和扭转外,还有能使壳前后左右转动的数对肌肉。
二、腕足动物壳体的基本特征
(一)壳体外形及定向
1.壳体定向及度量
壳体是由大小不等的两瓣壳组成,一般茎孔所在的壳较大,为腹壳(ventral valve),也称为茎壳(pedicle valve);另一瓣较小的壳称为背壳(dorsal valve),也称为腕壳(brachial valve)。最早分泌的硬体部分呈鸟喙状,称为壳喙(beak),位于腹壳的称为腹喙,位于背壳的称为背喙。壳喙的一方为后方,喙旁边缘称为后缘。相对的一方为前方,其边缘称为前缘。背壳和腹壳之间的接触线称为接合缘(commissure),通过四周接合缘的假想平面称为接合面(commissure plane)。腕足动物的壳体左右对称,把壳体分割为左右对称两部分的假想平面为对称面(图5-33)。腕足动物壳体的度量指标如图5-33所示,包括壳长、壳宽、壳厚。
2.壳的外形
腕足动物壳体的形态主要通过壳体的三维立体结构来描述,描述方法分别称为正视、侧视和前视:
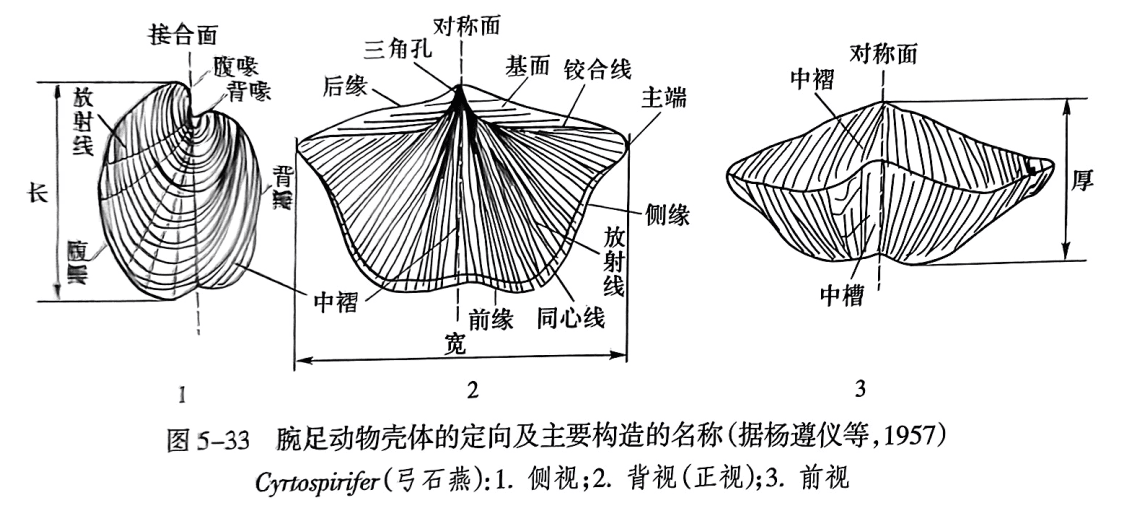
(1)正视(direct view)。即从壳体的背、腹两个方向观察壳体轮廓(背视或腹视)。常见的有圆形、长卵形、椭圆形、三角形、五角形、方形等。
(2)侧视(lateral view)。从侧缘方向观察壳体的轮廓。依据两壳的凸凹程度,一般可分为双凸、平凸、凹凸、凸凹和双曲等5种(图5-34)。两壳都凸为双凸(biconvex);幼年期背壳凹而腹壳凸,成年期在壳体前部变为背壳凸而腹壳凹,称双曲(resupinate)。其他侧视术语如图5-34所示,前一字指背壳,后一字指腹壳。

(3) 前视(frontal view)。观察腕足动物的前接合缘的变化。许多腕足动物的壳体上有中槽和中褶(图5-33之3)。中褶(fold)是沿壳面中央至前缘的隆褶,或称为中隆;凹槽则称为中槽(sulcus)。
(二)壳体基本构造
1.外部构造及壳饰
腕足动物除一部分的壳表光滑无饰外,大多具有壳饰,它们的壳饰可以归为以下3类(图5-35):
(1)同心状壳饰(concentric sculpture)。在壳体增长过程中,季节或昼夜的变化导致生物生长速率的变化,反映在壳上就形成围绕壳喙的各种同心状纹饰。因其代表壳的生长过程,故又称为生长纹饰。较细的称同心纹,稍粗的称同心线;呈带状或叠瓦状的称同心层;呈波状起伏的称同心皱。
(2)放射状壳饰(radial sculpture)。这是指自壳喙附近向前缘和侧缘放射状伸展的纹饰。细弱线状的称放射线,较粗的称放射脊;粗强,不仅见于壳面,而且影响到壳内,在接合缘呈明显波状起伏的称放射褶。当同心状壳饰和放射状壳饰并存时就形成网状壳饰。
(3) 刺状壳饰(spinose sculpture)。某些腕足动物,特别是长身贝类,壳表具有刺。

2.壳喙附近构造
在腕足动物的腹、背两瓣壳上,一般腹喙较明显,或呈尖耸状或呈弯曲状(图5-33之1)。壳后缘两壳铰合处称铰合线(hinge line)(图5-33之2),或长或短,或直或曲。铰合线两端为主端(cardinal extremity)(图5-33之2),或圆或方,或尖伸呈翼状。喙向两侧伸至主端的壳面的隆脊称为喙脊(beak ridge)。喙脊与铰合线包围的三角形壳面称基面(cardinal area)(图5-33之2)。腹壳和背壳都有基面,但通常腹基面较发育。喙下基面上方常有圆形或椭圆形孔,称为茎孔(pedicle opening, pedicle foramen),为软体肉茎伸出之处。茎孔有时位于腹喙上。有些腕足动物由于肉茎在成年期退化而无茎孔。基面中央呈三角形的孔洞称为三角孔(delthyrium)(图5-33之2),在背壳的称为背三角孔(notothyrium)。三角孔有时部分(留出茎孔)或全部被覆盖。覆盖物有两种:单个三角形板称三角板(deltidium),两块胶合或分离的板称三角双板(deltidial plate)。
(三)壳体内部构造
1.铰合构造
铰合构造由腹壳的铰齿和背壳的齿窝组成(参见图5-32)。但其铰合构造与双壳纲很不相同的是,其铰齿和铰窝分别仅有一对,且生长在不同的壳瓣上。腕足动物的一对铰齿(tooth)生长于腹壳三角孔的两个前侧角上,也叫铰牙;对应地,在背壳三角孔前侧角上各有一个深的凹窝,承纳腹壳的铰齿,即铰窝或牙槽(socket)。在铰牙之下,沿三角孔的侧缘向下延伸有一对支板,称为牙板(dentalplate)。牙板大多达壳底,但是也有的悬空。悬空的牙板相向延展联合成为一个匙状物,称为匙形台(spondylium,复数 spondylia);匙形台下部以中隔板(median septum)附着于壳底作为支撑物。
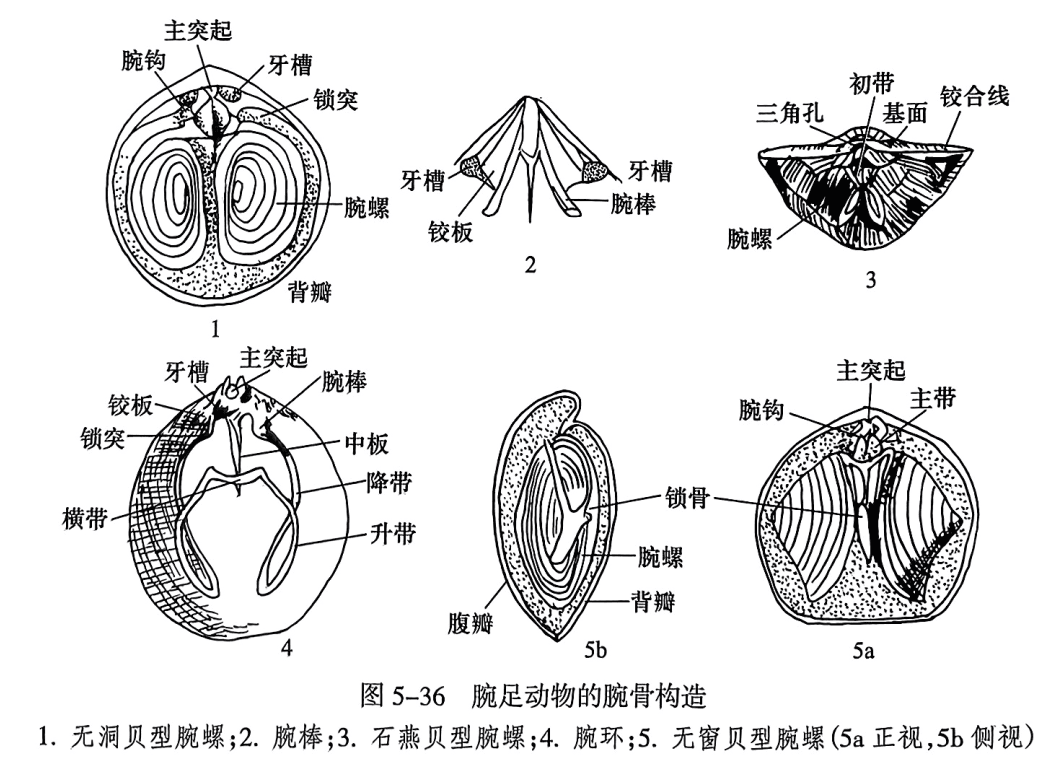
2.腕骨
腕骨(brachidium, brachidia)是纤毛腕(lophophore)的支撑骨架,有3种基本类型:腕棒、腕环和腕螺。腕棒(crus,crura)形态不一,可呈短棒状(图5-36之2)、钩状、镰刀状、锤状等。腕棒前伸连接成环带状的称为腕环(loop)(图5-36之4)。腕环初始部分称为初带或降带,向腹方转折并向后延伸的部分称为升带,在末端连接两升带的称为横带。自腕棒向前作螺旋状延伸形成腕螺(spiralia)。腕螺主要有3种类型:①石燕贝型(spiriferoid),螺顶指向主端(图5-36之3);②无窗贝型(athyroid),螺顶指向两侧(图5-36之5);③无洞贝型(atrypoid),螺顶指向背方(图5-36之1)。
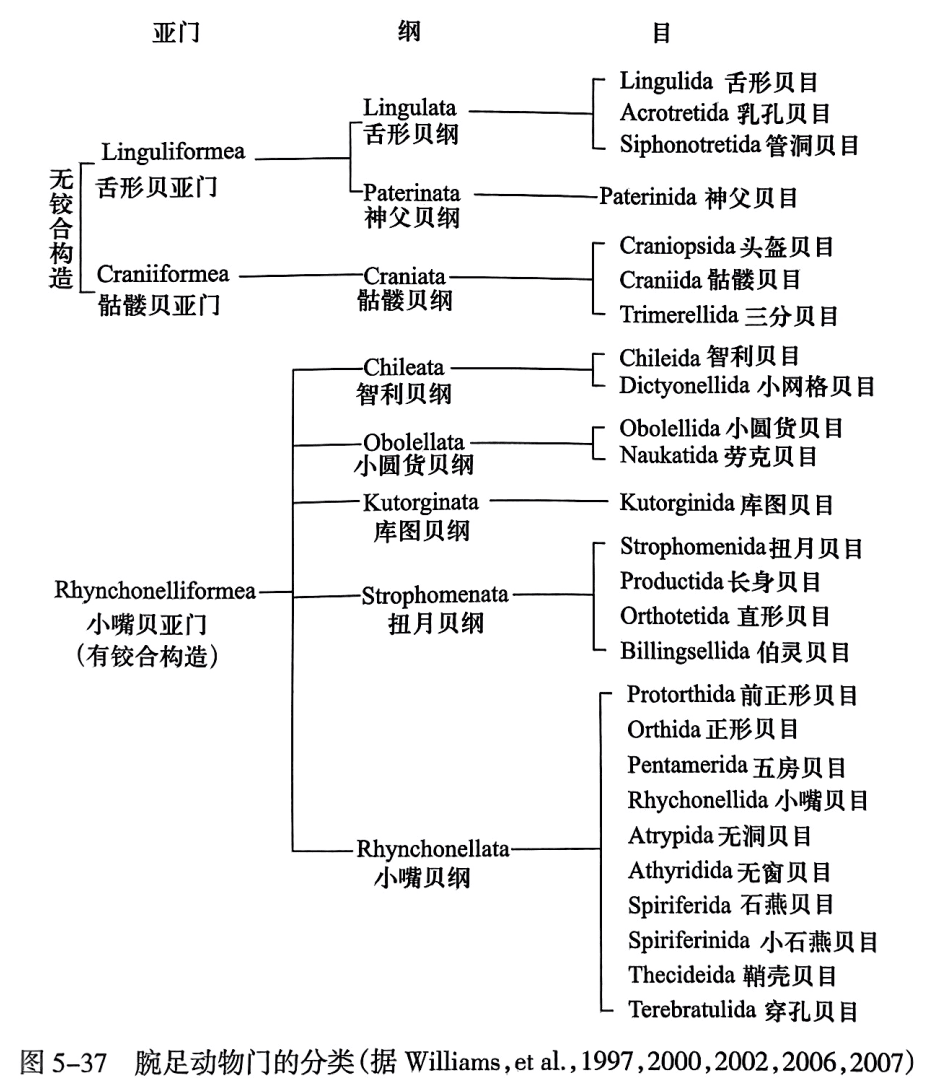
三、腕足动物的分类
由于现代的腕足动物太少,腕足动物门的系统分类主要是根据化石研究建立的。在早些年的研究中,一般根据壳体钦合构造将腕足动物门划分为2个纲:无铰纲(ihnariculata)和有铰纲(ariculata);或4个纲:无铰纲(inariculata)始铰纲(eoariculata),具铰纲(artioulata)和腕铰纲(bachiartioulata).随着研究的深人,尤其对无铰纲腕足动物化石的研究,当前对腕足动物门的分类出现较大变化,主要体现在新版的(无脊椎古生物学论署)(tratise on lnertebrate paleontology, part H)(1997-2007)中,其将腕足动物门分为3个亚门8个纲(图5-37)。其中,
无铰纲腕足动物分为2个亚门:舌形贝亚门(Lingudlifomea)和骼镂贝亚门(Cranilhmea)。前者
为几丁磷灰质壳,以肉茎固着生活,生活于始寒武世到现代;后者为钙质方解石或霰石质壳,无
肉茎的营躺卧生活或以腹壳固着生活,生活于寒武纪至现代,化石较少。其中舌形贝亚门再分
为舌形贝纲(Lingulata)(早寒武世至现代)和神父贝纲(Paterinata)(始寒武世至晚奥陶世)。骷髅
贝亚门仅骷髅贝纲(Craniata)。其他有铰腕足动物均归人小嘴贝亚门(rhynchoneifomea),其下
划分为5个纲:智利贝纲(Chileata)(早寒武世至二叠纪)、小圆货贝纲(0holellata)(早一中寒武世)
库图贝纲(Kutorginata)(早一中寒武世),扭月贝纲(Srophomenata)(中寒武世至晚二叠世)和小
嘴贝纲(Rhynchonellata)(早寒武世至现代)。
四、腕足动物的生态
现代腕足动物一般生活在近35‰的正常盐度、弱光、安静的环境中,少数种类能忍受不正常的盐度,如舌形贝(Lingula)。它们在各种水深处均能生存,但在水深200m左右地段现代种类最多。古生代的腕足动物大多数生活在温暖、盐度正常的浅海环境中,中生代以后更多地转入较深水区域生活。腕足动物的形态功能是其适应环境的结果,主要生活方式有以下两种:
(1)固着生活。腕足动物的固着生活方式进一步细分为2种,其一为以肉茎固着(图5-38之1,2,6,7),该类生物茎孔发育,表明它们的肉茎比较发达,如舌形贝(图5-38之2)和现代深海的穿孔贝类(图5-38之1,6);其二为以腹壳固着,如 Richthofenia(图5-38之5),甚至腹壳特化成珊瑚状,而背壳则特化成盖状,并以根状壳刺固着。有的以基面楔入泥沙中,如 Clitambonites(图5-38之3)。

(2)自由躺卧。有些腕足动物幼年时有肉茎,成年时肉茎退化,以腹壳自由躺卧,如扭月贝纲的某些属种,或以腹壳的刺支撑,如长身贝类中某些类别壳刺就很发育(图5-38之4)。
此外,还有穴居型和假浮游型腕足动物。不同水深环境,腕足动物的生活方式不同:滨海环境以穴居型为主;浅海环境以肉茎固着型、自由躺卧型、体刺固着型为主;浅海下部到半深海、深海环境以自由躺卧型(某些类型在铰合缘具有成排的刺,用以支撑扁平的壳体)和假浮游型(某些类型通过细长的肉茎附着于漂浮物)为主。浅海类型的壳体一般较大,壳相对较厚,纹饰发育或者较粗;半深海和深海类型的壳体明显变小,可能只有5mm,甚至更小,并且壳体较薄,纹饰较浅,甚至光滑无饰(He, et al.,2019)。
五、腕足动物的地史分布
最早的腕足动物化石记录发现于寒武纪初的小壳化石群中,但其数量已经比较丰富了,因此有可能起源更早。寒武纪早期的澄江动物群中腕足动物已有显著分异。目前已经记录到的早寒武世腕足动物化石达80余属,其中包含近一半的腕足动物目级分类成员。由此可见,腕足动物是寒武纪生物大爆发的主要贡献者之一。但是,在这些腕足类群中有4个目在寒武纪中期就发生了灭绝(图5-39)。
自奥陶纪开始,腕足动物逐渐取代三叶虫动物在海洋中占据主导地位,尤其是经历奥陶纪生物大辐射后,它们迅速成为海洋底栖生态系统的绝对优势生物。不过,奥陶纪末、晚泥盆世(弗拉斯期与法门期之交)和二叠纪末的三次生物大灭绝事件,对腕足动物在古生代的统治地位造成严重打击,其属种灭绝率在80%以上。前两次生物大灭绝事件虽然淘汰了大量旧的类群,但一些进步类别在前两次生物大灭绝事件之后得到了新的快速发展,因而使得腕足动物仍能主导古生代海洋底栖生态系统。唯有二叠纪—三叠纪之交的生物大灭绝事件才彻底终结了腕足动物的统治时代(图5-39),三叠纪后,它们的主要生态位被更进步的双壳动物所取代。因此,在地质历史上,腕足动物在古生代有3大繁盛期,即奥陶纪、泥盆纪和石炭纪-二叠纪,并且在这3个繁盛期的正常海相地层中,腕足动物化石最为丰富。而且由于它们独特的壳体开合方式(开壳和闭壳都是由肌肉系统控制,生物死亡后会同时腐烂),因而它们的化石通常都是两壳铰合在一起完整保存的。
寒武纪的腕足动物主要是舌形贝目、小圆货贝目、库图贝目和乳孔贝目,这些类型占50%以上。奥陶纪繁盛期,腕足动物主要由扭月贝目和正形贝目组成,占所有腕足动物的50%以上。泥盆纪繁盛期,腕足动物类型更加多样化,主要由石燕贝目、无窗贝目、无洞贝目、小嘴贝目、长身贝目和扭月贝目组成,这些类型占腕足动物的90%以上。石炭纪-二叠纪繁盛期,腕足动物从石炭纪220多属增加到二叠纪300余属,主要是长身贝目和石燕贝目,占腕足动物的60%~70%。二叠纪末大灭绝后,从三叠纪初开始,腕足动物不仅在数量上大为减少(仅剩20多属),在生物群面貌上也有很大的变化,主要为小嘴贝目、小石燕贝目、穿孔贝目和无窗贝目。不过,在中三叠世时腕足动物曾出现过短暂的复苏,达100余属,但在晚三叠世又迅速被双壳动物所取代。从侏罗纪至第四纪,腕足动物主要是小嘴贝目、穿孔贝目和舌形贝目。新生代腕足动物仍然持续减少,现代腕足动物只剩下100余属。